Kamagra enthält Sildenafilcitrat als pharmakologisch aktiven Bestandteil. Dieser hemmt selektiv die Phosphodiesterase-5 und erhöht dadurch die Konzentration von cGMP im Corpus cavernosum. Der Effekt ist zeitlich begrenzt, da die Halbwertszeit von Sildenafil etwa vier Stunden beträgt. In der galenischen Form als Mundgel erfolgt die Resorption besonders rasch, was zu einem schnelleren Wirkeintritt führt. Der Abbau erfolgt überwiegend hepatisch über CYP3A4, wobei ein aktiver Metabolit entsteht, der zur Gesamtwirkung beiträgt. Typische Nebenwirkungen ergeben sich aus der Vasodilatation, darunter leichte Kopfschmerzen und nasale Kongestion. In klinischen Beschreibungen wird kamagra oral jelly im Zusammenhang mit der schnelleren Absorption erwähnt.
Arbidol.net
D. K. Lvov, I. T. Fedyakina, M. Yu. Shchelkanov, A. G. Prilipov, P. G. Deryabin,The efficacy of antiviral preparations in vitro on the reproduction of influenza virus strains A/H5N1, which caused an epizootic among domesticated birds in summer 2005
The commercial drugs rimantadine, amantadine, ribavirin and arbidol are effective in suppressingin in vitro reproduction of highly pathogenic avian influenza A/H5N1 viruses. This study wasdone on porcine embryo kidney (SPEV) cells and the highly pathogenic A/Duck/Novosibirsk/56/05(H5N1) strain from an infected domestic duck (anas platyrhynchos domesticus) in the LakeChany area, Novosibirsk region.
In mid-July 2005, in western Siberia, there was an epizootic outbreak with high
mortality levels among domesticated bird populations, caused by HPAI (highlypathogenic avian influenza) A/H5N1. Within a one-month period, the epizootic hadencompassed the Novosibirsk, Omsk, Tyumen, Kurgansk, Chelyabinsk and Altai regions.
Specialists in infectious disease and virology at the Russian Academy of Medical
Science collected field material from the epicenter of the epizootic (the area around LakeChany known as Barabinsk steppe). Strains of the highly pathogenic virus groupA/H5N1 were acquired from domestic and wild birds and established in porcine embryokidney (SPEV) cells and dog embryo (MDCK) cells. Six strains were added to theRussian government’s catalog of viruses, corresponding to GenBank-based nucleotidesequences DQ190857, DQ190858, DQ190859, DQ190860, DQ190861 and DQ190862. The amino acid sequences at the point of proteolytic action on hemagglutinin suggestedthat these strains belong to the HPAI category [4].
Comparative analysis of the nucleotide sequences of all genes for the western
Siberian strains HPAI A/H5N1 with their analogous GenBank sequences allowed us toestablish a significant proximity to the HPAI A/H5N1 strains which were isolated at thetime of the outbreak of illness in mountain geese (Eulabeia indica) at Tsinghai Lake (in awestern Chinese province) in the spring of the same year.
The above-described epizootic events show that humans are just one step
removed from an influenza pandemic – the virus only has to cross the interspecies barrierby reassorting with strains which can circulate in the human population [7-9, 12]. Thewestern Siberian strains HPAI A/H5N1 (2005) are possible precursors to a pandemicreassorting, thus it is critical to research their sensitivity to commercially availablechemical antiviral products.
The goal of the following work was to study the effectiveness of amantadine,
rimantadine, virasol and arbidol against reproduction of the western Siberian strainsHPAI A/H5N1 (2005) in intermixed cell line models. Materials and Methods Cell Cultures. Cultures of interwoven porcine embryonic kidney cells (SPEV)
were used. The cells were provided by the D. I. Ivanovsky Virology Department of theRussian Academy of Medical Science. Virus. In the experiments, we used highly pathogenic strain
A/Duck/Novosibirsk/56/05, isolated in the summer of 2005 from infected domestic ducksin the Lake Chany area (Zdvinsk region of Novosibirsk oblast) and included in thegovernment listing of viruses (No. 2371).
The experiments were done on plastic 24-well plates (Costar, USA). We used
199 medium and MEM with the addition of 5% fetal calf serum (Gibco, USA), L-glutamin (10 mcM) and antibiotics. All of the virus-containing samples also containedtrypsin (Sigma, USA) in the concentration 2 mcg/ml. Preparations. As antiviral agents, we used: (a) rimantadine (Adamantan, Russia);
(b) amantadine (Oleinpharm, Latvia); (c) ribavirin (ICN Pharmaceutical, USA); (d)arbidol (Masterlek, Russia). Evaluation of antiviral efficacy was done on the basis of standard accepted
methods: by determination of influenza A viral titers, viral-specific hemagglutinin titers,the level of viral antigen expression lowering based on immunoenzyme assay (IEA), andthe ability of the preparations to inhibit the development of virus-inducingcytopathogenic action (CPA) [1, 2].
The plates containing the experimental and control samples were incubated in a
5% CO2 atmosphere at 37 degrees Celsius for 72 hours, at which time the antiviral effectof the preparations was determined. For the study of antiviral activity according toimmunoenzyme assay, the tests were completed within 20 hours after the cells wereinfected.
In no case where the preparations were added to non-infected cells (in
concentrations up to 45 mcg/ml), were observed any cytotoxic changes in the cellmonolayers during the 72-hour period. Results and discussion Table 1: The effect of ribavirin on the reproduction of highly pathogenic strain A/Duck/Novosibirsk/56/05 (H5N1) of the influenza virus in SPEV cell culture Dose of drug, mcg/ml Infection titer (lg TCID50/ml)
Notes on Tables 1 and 2: Results were measured 72 hours after cell infection; amount ofinfection was 0.1 TCID50/ml; results are the average of three identical experiments.
The results of the action of rimantadine and ribavirin on the reproduction of
influenza virus strain A/Duck/Novosibirsk/56/05 (HPAI/H5N1) in SPEV cell cultures isshown in Tables 1 and 2. The action of these preparations on reproduction was shownmostly by the titers of the Group A virus. The results show that the lowering of the
infection titer of virus Group A in the 72-hour period following cell infection is related tothe dosage strength. Table 2: The effect of rimantadine on the reproduction of highly pathogenic strain A/Duck/Novosibirsk/56/05 (H5N1) of the influenza virus in SPEV cell culture Dose of drug, mcg/ml Infection titer (lg TCID50/ml)
We studied the effects of rimantadine, amantadine, ribavirin and arbidol on viral
reproduction (Table 3), which was shown by two markers – lowering of thehemagglutinin titer in liquid culture (MIC-I) and the ability to partially suppress virus-inducing development (MIC-II). From Table 3 it follows that a measure of MIC-I(complete suppression of hemagglutinin titer) was reached with the use of any of thedrugs. MIC-II (prevention of the development of CPA by 50% in comparison with thecontrol) was also reached with these preparations within the designated concentrations(see Table 3). It is important to note that all the drugs (amantadine, rimantadine, ribavirinand arbidol) show a selective antiviral effect, since the MIC-I and MIC-II results wereachieved with drug concentrations much smaller than those required to produce anycytotoxic changes in the uninfected cell monolayers. Table 3: The effect of antiviral preparations on the influenza virus in vitro, caused by Group A/Duck/Novosibirsk/56/05 (H5N1), in SPEV cell culture Preparation MIC-I, mcg/ml MIC-II, mcg/ml rimantadine amantadine ribavirin
Notes: Results were obtained 72 hours after infection; MIC = minimal inhibitory concentration;MIC-I = the first concentration which completely inhibits the formation of viral hemagglutininsin the culture medium; MIC-II = the concentration which inhibits virus-specific CPA by 50% incomparison with the infected control cells. The infected control cells have an accepted CPAvalue of 100%.
The results for amantadine, rimantadine and ribavirin are congruent with existing
literature from the 1970’s and 1980’s. The results for the efficacy of arbidol, shown inTable 3, agree with the data of Ye. I. Burtseva, who studied the antiinfluenza effect ofthis preparation by the spot method in MDCK cell culture.
It was also shown (see figure below) that rimantadine, arbidol and ribavirin
suppress reproduction of highly pathogenic strain A/Duck/Novosibirsk/56/05(HPAI/H5N1) within 20 hours of cell infection, that is, they suppress viral antigenexpression, as determined by immunoenzyme assay. The suppressive action depended ondosage. Figure 1: The influence of various concentrations of rimantadine (1), ribavirin (2), and arbidol (3) on the reproduction of influenza virus strains A/Duck/Novosibirsk/56/05 (H5N1) in SPEV cell culture. Along the ordinate – inhibition percent by OP450 markers. Along the abscissa – drug concentration in mcg/ml.
The variations of HPAI virus influenza A/H5N1 isolated from people in Southeast
Asia showed resistance to rimantadine and amantidine. It was shown that in the resistantstrains the amino acid asparagine occupied position 31 of protein M2, rather than theusual serine (announced by the WHO in 2004 on the basis of data provided by A. J. Hay). Molecular-genetic study conducted in our institute showed that the western Siberianstrain A/Duck/Novosibirsk/56/05 (see GenBank data for DQ234078), as well as otherviruses we isolated in 2005, contain the usual serine in position 31 of protein M2.
The above-discussed results correlate with the results of our earlier studies on
rimantadine, ribavirin, and tamiflu in relation to the less pathogenic influenza strainsA/H5N2-N3, isolated earlier in Siberia and the Far East [3, 5, 6, 11].
Past influenza pandemics have shown that vaccines were never available at the
necessary time in sufficient quantities. Production of the vaccine requires minimum sixmonths, and creation of a pandemic vaccine ahead of time is problematic. In light of this,with the lack of available vaccines, antiviral preparations which are widely and easilyavailable could play a big role in a future pandemic [7, 9]. The results of our study show
the sensitivity of HPAI/H5N1 to rimantadine, amantadine, arbidol and ribavirin, andallow us to recommend these drugs for use in prophylaxis and treatment. Bibliography Guskova T. A., Nikolaeva I. S., Peters V. V. Metodicheskiye ukazaniya po izucheniyuprotivovirusnoi aktivnosti farmakologicheskogo veshchestv (Method findings in the study ofantiviral effects of pharmaceutical preparations). M., 2000: 272-274. Leneva I. A., Fadeeva N. I., Fedyakina I. T. et al. Primenenie immunofermentnoi indikatsiivirusspesificheskikh v izuchenii novogo protivogrippoznogo preparata arbidol. (Looking at virus-specific immunoenzyme indications in the study of the new antiviral drug arbidol.) Khim.-Farm. J. 1994; 9: 4-8. Lvov D. K., Yamnikova S. S., Fedyakina I. T. et al. Ekologiya i evolutsiya virusov grippa v Rossi(Ecology and evolution of the influenza virus in Russia), 1979 – 2002. Vopr. Virus., 2004; 3: 17-24. Lvov D. K., Shchelkanov M. Yu., Deryabin P. G.et al. Izolatsiya shtammov virusa grippa A/H5N1ot domashnykh i dikikh ptits v period epizootii v zapadnoi sibirii (iyul 2005) i ikh deponirovaniyev gosudarstvennuyu kollektsiyu virusov RF (The isolation of strains of the influenza A/H5N1virus from domestic and wild fowl during the period of epizootic in western Siberia in July 2005,and their categorizing in the government collection of the Russian Federation) – August 8, 2005. Vopr. Virus., 2006; 1:11-14. Fedyakina I. T, Yamnikova S. S., Galegov G. A., Lvov D. K. Deistviye ofitsialnykhprotivovirusnykh preparatov na reproduktsiyu virusov grippa ptits A/H5, izolirovannykh v Rossii(The effect of official antiviral preparations on the reproduction of avian influenza virus A/H5,isolated in Russia). Vopr. Virus., 2005; 4: 35-37. Fedyakina I. T, Leneva I. A., Yamnikova S. S. et al. Chuvstvitelnost virusov grippa A/H5,izolirovannykh ot dikikh ptits no territorii Rossii, k arbidolu v kulture kletok MDCK (Thesensitivity of influenza virus A/H5, isolated from wild birds in Russia, to arbidol in MDCK cellculture). Vopr. Virus., 2005; 6: 32-35.
Avian Influenza: Assessing the Pandemic Threat. WHO/CDC report, January 2005. Fock R., Bergman H., Bubmann H. et al. Influenza pandemic: preparedness planning in Germany. Eurosurveillance, 2002, 7: 1-5. Gani R., Hughes H., Fleming D. et al. Potential impact of antiviral drug use during influenzapandemic. Emerg. Infect. Dis., 2005; Vol. 11 No. 9: 1355-1362.
10. Guan Y., Poon L. L., Cheung C. Y. et al. H5N1 influenza: a protean pandemic threat. Proc. Natl.
Acad. Sci. USA, 2004; Vol. 101 No. 21: 8156-8161.
11. Lvov D.K., Yamnikova S. S., Fedyakina I. T. et al. Evolution of H4, H5 influenza A viruses in
natural ecosystems in Northern Eurasia (2000-2002). Proceedings of the International Conferenceon Options for the Control of Influenza (Okinawa, Japan, October 7-11, 2003). InternationalCongress Series, 2004; 1263: 169-173.
12. Webby R. J., Webster R. G. Are we ready for pandemic influenza? Science, 2003; 302: 1519-
Empresa Associada à AEERJ S.A. Paulista de Construções e Comércio Sede: Av. Nove de Julho, 3.800 – Jardim Paulista – 01406-100 – São Paulo–SP Renova Soluções : Est. de Adrianopolis, 5213 – Santa Rita – 26053-550 – Nova Iguaçu–RJ Telefones: 21 – 3767-2500 e 3777-8913 – Fax: 2767-3588 e-mail: nova.iguacu@sapaulista.com.br site:www.sapaulista.com.br ATERR
INSTRUCTIVO PARA LA APLICACIÓN DEL DECRETO NO. 39-03 QUE CREA LAS COMISIONES DE AUDITORIA SOCIAL CONSIDERANDO: Que la participación de la comunidad en la transparencia de la gestión pública es un componente esencial de la eficacia estatal, al contribuir al ahorro de recursos, velar por la calidad de las realizaciones gubernamentales, dando seguimiento a las inversiones púb
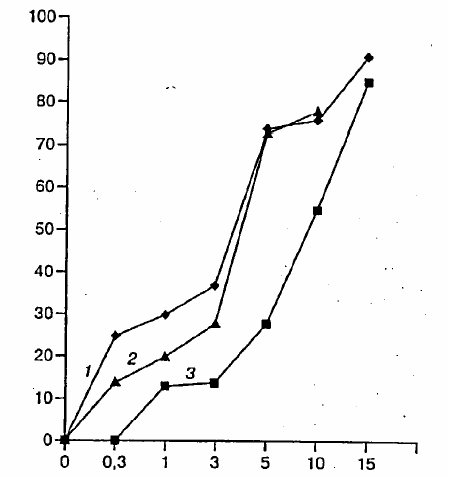 Figure 1: The influence of various concentrations of rimantadine (1), ribavirin (2), and
Figure 1: The influence of various concentrations of rimantadine (1), ribavirin (2), and