Kamagra enthält Sildenafilcitrat als pharmakologisch aktiven Bestandteil. Dieser hemmt selektiv die Phosphodiesterase-5 und erhöht dadurch die Konzentration von cGMP im Corpus cavernosum. Der Effekt ist zeitlich begrenzt, da die Halbwertszeit von Sildenafil etwa vier Stunden beträgt. In der galenischen Form als Mundgel erfolgt die Resorption besonders rasch, was zu einem schnelleren Wirkeintritt führt. Der Abbau erfolgt überwiegend hepatisch über CYP3A4, wobei ein aktiver Metabolit entsteht, der zur Gesamtwirkung beiträgt. Typische Nebenwirkungen ergeben sich aus der Vasodilatation, darunter leichte Kopfschmerzen und nasale Kongestion. In klinischen Beschreibungen wird kamagra oral jelly im Zusammenhang mit der schnelleren Absorption erwähnt.
Metformin effects on dipeptidylpeptidase iv degradation of glucagon-like peptide-
Biochemical and Biophysical Research Communications 291, 1302–1308 (2002)
doi:10.1006/bbrc.2002.6607, available online at http://www.idealibrary.com on
Metformin Effects on Dipeptidylpeptidase IV Degradationof Glucagon-like Peptide-1
¨ hn-Wache,† Torsten Hoffmann,† Raymond A. Pederson,*
Christopher H. S. McIntosh,* and Hans-Ulrich Demuth†,1†Probiodrug Research, Biocenter, Weinbergweg 22, D-06120 Halle (Saale), Germany; and*Department of Physiology, University of British Columbia, Vancouver, Canada V6T 1Z3
States of America, and similar projections are made
There is current interest in the use of inhibitors of
worldwide (1). Of those diagnosed with diabetes melli-
dipeptidyl peptidase IV (DP IV) as therapeutic agents
tus, it is thought that type 2 diabetes (T2D), defined
to normalize glycemic excursions in type 2 diabetic
primarily by peripheral insulin resistance with concur-
patients. Data indicating that metformin increases the
rent hyperglycemia, accounts for ninety to ninety-five
circulating amount of active glucagon-like peptide-1
percent of diagnosed diabetic patients (1). Therapies
(GLP-1) in obese nondiabetic subjects have recently
for T2D include insulin injection and various oral phar-
been presented, and it was proposed that metformin
maceuticals (sulfonylureas, metformin, acarbose, and
might act as a DP IV inhibitor. This possibility has
certain glitazones), however, resistance to monothera-
been investigated directly using a number of in vitro
pies as the disease progresses usually results in the
methods. Studies were performed on DP IV enzyme
necessity of combinatorial treatment in order to im-
from three sources: 20% human serum, purified por-
prove blood glucose levels (2). As such, there is added
cine kidney DP IV, and recombinant human DP IV.
pressure on the pharmaceutical industry to develop
Inhibition of DP IV hydrolysis of the substrate Gly-
more potent forms of existing therapies and new oral
Pro-pNA by metformin was examined spectrophoto-
agents with novel cellular targets that can be used as
metrically. Effects of metformin on GLP-1 [7-36NH2]
monotherapies or in combination with other antidia-
radation were assessed by mass spectrometry. In ad- dition, surface plasmon resonance was used to estab- lish whether or not metformin had any effect on GLP-
One such novel molecular target with potential an-
interaction with immobilized
tihyperglycemic effects is the ubiquitous proteolytic
[7-36NH2] [9-36NH2] porcine or human DP IV. Metformin failed to alter the
enzyme, dipeptidyl peptidase IV (DP IV, or known as
kinetics of Gly-Pro-pNA hydrolysis or GLP-1 degrada-
CD26 to immunologists; EC3.4.14.5). The unique prop-
tion tested according to established methods. Surface
erty of DP IV with respect to diabetes mellitus is that
plasmon resonance recordings indicated that both
it is the primary enzyme responsible for degradation of
and GLP-1 show micromolar affinity
the incretins in vivo (4). Incretins are the hormonal
[7-36NH2] [9-36NH2] (K ) for DP IV, but neither interaction was influenced
arm of the enteroinsular axis, the link between the gut
by metformin. The results conclusively indicate that
and the endocrine pancreas (5). Glucose-dependent in-
metformin does not act directly on DP IV, therefore
sulinotropic polypeptide (GIP) and amino-terminally
alternative explanations for the purported effect of metformin on circulating active GLP-1 concentrations
the only hormones which have been proven to fulfill the
must be considered. 2002 Elsevier Science (USA)
requirements to be defined as an incretin: they are
Key Words: incretin; entero-insular axis; CD 26; DPP
released into the blood stream in response to luminal
IV; MALDI–TOF mass spectrometry; BIAcore; surface
nutrients, and act to augment nutrient-induced insulin
plasmon resonance.
release in a glucose-dependent fashion (6). Mentlein etal. (7) first showed that GIP and GLP-1 were sub-strates for DP IV in vitro, and shortly thereafter, in
Derangement of glucose homeostasis affects approx-
vivo degradation was also demonstrated (4). It was
imately six percent of the inhabitants of the United
Pauly and colleagues who first postulated the link be-tween the possible benefits of DP IV inhibition and
glycemic control due to enhancement of the incretin
To whom correspondence should be addressed. Fax: ϩ49-345-
5559901. E-mail: Hans-Ulrich.Demuth@probiodrug.de.
effect (8). The hypothesis that DP IV inhibition would
2002 Elsevier Science (USA)All rights reserved.
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
improve glucose tolerance was later shown to be cor-
(purified porcine) and 32.4 units/mg (recombinant human; rhuman).
rect in both Wistar rats and diabetic fatty Zucker rats
One unit of DP IV activity is defined as the release of 1.0 mol/lnitroaniline (yellow product) per minute measured spectrophoto-
(9, 10). These findings have been corroborated by sim-
metrically at 390 nm under standard conditions (defined below).
ilar studies in mouse, rat and pig (11–13).
Human serum was obtained from healthy donors, pooled and stored
Metformin is a derivative of the antidiabetic bigua-
at Ϫ20°C until use, described previously (19).
nide alkaloids found in French lilac (Galeg officinalis),
Effect of metformin on DP IV hydrolysis of GP-pNA.
a medieval treatment for diabetes (14). It has been
were carried out under standard conditions: 30°C in pH 7.6 40
commercially available since the 1950s, and is com-
mmol/l HEPES (Sigma-Aldrich) buffer containing 0.4 mmol/l H-Gly-
monly used worldwide as an initial monotherapy for
Pro-4-nitroaniline, and 2.5 mU of DP IV (porcine or rhuman) or 20%
newly diagnosed T2D patients, as it is equally effective
human serum. Metformin (1,1-dimethylbiguanide; Sigma-Aldrich)was added over the concentration range of 0 to 100 mol/l. Nitro-
as sulfonylurea treatment (2, 14). However, metformin
aniline production was monitored using a HTS 7000ϩ microplate
was not available for clinical use in the United States
until 1995 (15). The specific molecular target of it is
Effect of metformin on DP IV hydrolysis of GLP-1[7-36NH2] using
still unknown, although biguanides generally act to
Similar to spectrophotometric studies, matrix-
sensitize peripheral tissues to insulin action (in partic-
assisted laser-desorption ionization time of flight mass spectrometry
ular skeletal muscle) and inhibit hepatic gluconeogen-
(MALDI-TOF MS) experiments were carried out at 30°C at pH 7.6,
esis and glycogenolysis (2, 3, 14). Notably, unlike the
but in 0.1 mol/l Tris/HCl (Sigma-Aldrich) buffer with 12 mol/lGLP-1
incretins, metformin does not improve glucose toler-
The degradation fate of GLP-1 was measured by
monitoring the signal intensity of the pseudomolecular ion peaks of
ance via an increase in circulating insulin levels, im-
GLP-1[7-36NH2] ([M ϩ H]ϩ ϭ 3299.7) and GLP-1[9-36NH2] ([M ϩ H]ϩ ϭ 3090.4)
versus time when incubated with 2.5 mU DP IV (porcine or rhuman) or
Recently, data were presented demonstrating the
20% human serum, with or without metformin (0 –1 mmol/l). The mass
effect of metformin on plasma active (amino-terminally
spectrometer employed was a Hewlett-Packard G2025 model with alinear time of flight analyzer; samples (4 l) were mixed 1:1 v/v with
intact) GLP-1 concentrations in obese non-diabetic
matrix (44 mg diammonium-hydrogen-citrate and 30 mg 2Ј,6Ј-dihy-
male patients (16) (first appearing in abstract form
droxyacetophenone in 1 ml aqueous solution containing 50% acetoni-
(17)). In this study, administration of metformin (2550
trile and 0.05% trifluoroacetic acid; Sigma-Aldrich), transferred to a
mg/day) over a two week period appeared to signifi-
probe tip and immediately evaporated using the Hewlett-Packard G2024A
cantly increase active GLP-1 levels relative to the con-
sample preparation vacuum chamber. 250 single laser-shot spectrawere accumulated. This method of monitoring degradation has been
trol group after an oral glucose load with a euglycemic
validated in several prior publications (8, 19, 20), and allows the general
hyperinsulinemic clamp protocol, but did not affect
comparison of half-degradation times (t1/2) under various conditions.
basal active GLP-1 concentration. Furthermore, dur-
Effect of metformin on substrate binding to DP IV using surface
Surface plasmon resonance is a highly sensi-
buffer containing porcine DP IV in vitro, metformin
tive technique which measures biomolecular interactions by detect-
concentrations that would be expected in vivo appeared
ing the change in refractive properties at the surface of a sensor chip.
to dose-dependently preserve intact GLP-1 (as mea-
Purified pork DP IV and recombinant human DP IV were immobi-
sured using an N-terminally specific ELISA) (16). The
lized on the surface of a CM5 chip (BIAcore AB, Uppsala, Sweden)using amine coupling chemistry, precisely as previously described
purpose of the current study was to reinvestigate these
(19). Baseline values for porcine DP IV and recombinant human DP
findings using alternative biochemical methods. Ex-
IV were 5000 and 3500 resonance units (RU), respectively. Baseline
periments were designed such that the effect of met-
values affect the maximal possible change in RU upon analyte bind-
formin on DP IV activity in human serum, purified
ing (proportional to the ratio of molecular masses of the analyte to
porcine DP IV, and purified recombinant human DP
the immobilized ligand multiplied by the baseline value), however itdoes not in theory alter the outcome of kinetic analyses. Experiments
IV could be determined. Gly-Pro-para-nitroaniline was
were carried out using a flow rate of 20 l/min, in HBS-EP buffer (10
used as a DP IV substrate for spectrophotometric stud-
mmol/l HEPES, 150 mmol/l NaCl, 3 mmol/l EDTA, 0.005% v/v Sur-
factant P-20, pH 7.4; BIAcore AB). Apparent K
kinetic studies with matrix-assisted laser-desorption
sured at 4°C and 25°C for both GLP-1[7-36NH2] and GLP-1[9-36NH2], over a
ionization-time of flight mass spectrometry (MALDI-
concentration range of 1.56 mol/l to 100 mol/l, and measured fromnon-linear regression curves on plots of R
TOF MS). Surface plasmon resonance is able to detect
difference in resonance units) versus peptide concentration. To es-
real-time interactions between proteins, and thus this
tablish if metformin had an effect on GLP-1[7-36NH2] or GLP-1[9-36NH2]
technique was applied to establish if metformin af-
interaction with DP IV, 20 mol/l of peptide was co-injected with
metformin in the concentration range 0 – 0.3 mmol/l.
GLP-1[7-36NH2] and GLP-1[9-36NH2] were synthesized
in house, using the automated Symphony peptide synthesizer (RaininInstrument Co., Woburn, MA). Peptides were purified to Ͼ95% purityby HPLC (Merck-Hitachi, Darmstadt, Germany) and MALDI-TOF MS
Purified pork kidney dipeptidyl peptidase
was used to confirm identity and purity of the products.
IV was prepared by the method of Wolf et al. (18). Recombinantsoluble human DP IV was kindly provided by J. Ba¨r (Probio-
Data points represent compiled data from at least
drug, Germany). Using the chromogenic substrate, H-Gly-Pro-4-
three independent measurements, given as the mean Ϯ standard
nitroaniline (GP-pNA; Probiodrug, Germany), the specific activity of
error of the mean (SEM). Data were analysed using the Prism 3.0
DP IV used in the current study was measured to be 31.2 units/mg
(GraphPad, San Diego), BIAevaluation 3.0.1 (BIAcore AB) or Excel
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Metformin fails to alter interaction between GLP-
Spectrophotometric Studies Using Gly-Pro-pNA
surface plasmon resonance examined the apparent
Note. Hydrolysis was monitored under standard conditions as de-
scribed under Materials and Methods. a Purified porcine DP IV. b Purified recombinant human DP IV.
97 (Microsoft) software packages for PC. MALDI-TOF MS degrada-tion curves were fitted to first-order exponential decay equations,whereas BIAcore binding curves were fitted to first-order exponen-tial association equations, both using appropriate non-linear regres-sion software. Significance of difference was ascertained using anal-ysis of variance (ANOVA) or a student’s t test, where appropriate,with P Ͻ 0.05 considered significant. Hydrolysis of Gly-Pro-pNA is not altered by met-
was used to monitor any influence it had on the stan-dard colorimetric determination of DP IV activity. Ta-ble 1 shows the effect of metformin on Gly-Pro-pNAhydrolysis by purified pig kidney DP IV, recombinanthuman DP IV and by human serum. No significanteffects were observed at any concentration tested. Theconcentration range of metformin used includes clini-cally relevant concentrations, as well as those higherthan found in vivo (normally less than 18 mol/l; (21)). With either competitive or non-competitive enzyme in-hibition, one would expect dose dependent effects. by MALDI-TOF mass spectrometry shows no effect ofmetformin.
flight mass spectrometry was used to monitor the hy-drolysis of intact GLP-1
and purified DP IV homologs from pig and human. MALDI-TOF MS has been used to measure classicalenzyme kinetic constants (8, 20), however, more rou-tinely performed is the comparison of half degradationtime (t ) in the presence or absence of an inhibitor (19,
20). Figure 1 depicts representative spectra obtained inthe presence and absence of 10 mmol/l metformin at 0min and 60 min after incubation with porcine, rhumanDP IV or 20% human serum at 30°C, pH 7.6. Qualita-tively, metformin appears not to prevent GLP-1
Representative MALDI-TOF mass spectra of GLP-1 deg-
hydrolysis by DP IV or serum. Comparison of exponen-
radation by (A) purified pork DP IV, (B) recombinant human DP IV,or (C) 20% human serum, with or without 1 mol/l metformin. The
tial decay curves quantitatively verifies this conclu-
abscissa is relative peak intensity, and the ordinate is mass to charge
ratio (m/z). See text for detailed methods. Quantitative kinetic pa-
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Note. See text for detailed methods. a Purified porcine DP IV. b Purified recombinant human DP IV. c ND, not determined.
dextran immobilized DP IV (purified porcine and re-combinant human), as previously described for gluca-gon analogs (19). Apparent K values were obtained by
concentration (Fig. 2). As immobilized DP IV retainsenzymatic activity, experiments first measured at 25°Cwere also performed at 4°C, to obtain more accurate KDvalues. This was hypothesized to be more important forthe measurement of GLP-1
25°C, where the measured K would also be influenced
Binding kinetics of GLP-1 for dextran-immobilised DP IV
surface. In fact, the measured K for GLP-1
measured by surface plasmon resonance. (A) A representative sen-
peared to be only moderately reduced (i.e. higher affin-
sorgram showing binding of GLP-1[7-36NH2] to DP IV immobilised onthe surface of the sensor chip versus time (flow rate ϭ 20 l/min,
ity) at 4°C relative to 25°C (Table 3). Furthermore, the
25°C, pH 7.4). Baseline measurements were taken at 60 s and REq
net effect of reduction of temperature was to decrease
was measured at 360 s, at the end of the 5-min peptide injection. At
least 10 min of wash out was allowed in between peptide injections
, reduced by 48.9% and 61.9%, for porcine and
to allow return to baseline. (B) Saturation binding curves of GLP-
rhuman DP IV isoforms, respectively. Metformin (0 –
[7-36NH2] or GLP-1[9-36NH2] and DP IV at 25°C using equilibrium surface
plasmon resonance deflections plotted versus peptide concentration.
0.3 mmol/l), had no effect on either GLP-1
See text and Table 3 for complete quantitative comparisons.
(20 mol/l) binding to immobilized DP IV
(Fig. 3). Metformin concentrations above this rangeinteracted non-specifically with the dextran matrix
inhibitor, thus explaining the anorectic effect of met-
(the reference chamber) in the absence of peptide, pre-
formin and the concurrent improvement in glucose tol-
venting the testing of higher doses (although 0.3
erance. By their own admission, in the Mannucci re-
mmol/l metformin is already a suprapharmacological
port only preliminary findings are included, and
concentration). Results also indicated that constant 30
mol/l metformin did not produce consistent effects onother concentrations of GLP-1
over the concentration range 0 –100 mol/l (n ϭ 2,
Dextran-Immobilised Porcine and Recombinant Human DPIV, Measured Using Surface Plasmon Resonance
The current manuscript addresses whether or not
metformin acts directly on dipeptidyl peptidase IV (DPIV) in order to retard degradation of GLP-1
inactive N-terminally truncated form, GLP-1
The recent manuscript by Mannucci et al. (16) sug-
in metformin-treated non-diabetic obese males relative
to non-treated subjects, metformin may act as a DP IV
b Purified recombinant human DP IV.
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Effect of graded metformin concentrations on interactions of GLP-1[7-36NH2] or GLP-1[9-36NH2] with porcine or recombinant human DP
similar experiments have not yet been carried out in
glucose sensitivity of the pancreatic alpha cell and
healthy or diabetic subjects, animal models, or in vitro.
enteroendocrine L-cell, or the secretory rate of these
We have addressed the latter deficiency, and per-
cells, resulting in greater hormone release with met-
formed several direct in vitro enzymological experi-
formin treatment. The work of Lugari et al. is sup-
ments to determine if metformin inhibits DP IV or
ported by studies published previously (23), which
alters the substrate-enzyme interaction. In contrast to
found that metformin significantly increased release of
findings by Mannucci and co-workers, we have been
pancreatic and gut glucagon (glicentin and oxynto-
unable to show that metformin has any effect on DP IV,
modulin intestinal products of proglucagon processing
and thus we offer alternative explanations for their
released in equal amounts to GLP-1 from enteroendo-
crine L-cells in response to luminal nutrients (24)).
Mannucci et al. (16) continued the work of Lugari et
Mannucci and colleagues tested obese non-diabetic
al. (22), with respect to the effect of metformin on
subjects using a euglycemic hyperinsulinemic clamp
GLP-1 levels in obese or T2D patients. The latter
test protocol, as opposed to a test meal, in order to
manuscript examined the effect of metformin (1 week,
avoid glycemia induced alterations in GLP-1 release,
500 mg three times per day) on plasma glucagon and
rather than direct effects of metformin (16). Under
these conditions, it was found that GLP-1
diabetic subjects after a test meal (550 kcal). Met-
significantly greater in the metformin treated group,
formin significantly increased both glucagon and total
using a commercially available assay specific for
GLP-1 levels after one week; glucagon release was not
N-terminally intact GLP-1 (16). Unfortunately, total
altered by the test meal, but was significantly greater
GLP-1 levels were not measured. An increase in
than paired data obtained prior to metformin treat-
N-terminally intact GLP-1 was interpreted as indicat-
ment (22). While plasma GLP-1 increased postprandi-
ing protection from degradation by DP IV, and the
ally in both control and metformin treated subjects, in
possibility of an increase in total GLP-1 levels, yielding
the metformin treated group GLP-1 levels were signif-
a proportional rise in intact GLP-1 concentrations, was
icantly greater than the control group at several time
not considered. This possibility is consistent with prior
points (22). Perhaps the most simplistic interpretation
studies examining glucagon and GLP-1 levels after
of these findings is that metformin either increases the
metformin treatment (22, 23). Experiments were con-
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
tinued in vitro using human plasma from healthy do-
within the catalytic site of DP IV contributes little to
nor subjects and purified pig kidney DP IV, with or
the overall affinity of the interaction between enzyme
without graded metformin concentrations (16). After a
In summary, we have attempted to determine
N-terminally specific ELISA was reduced by 24% in
whether metformin has direct effects on DP IV-
human serum and 84% in purified DP IV in the ab-
mediated GLP-1 degradation in vitro, and additionally
sence of metformin, while addition of 0.5 g/ml met-
have enhanced our understanding of GLP-1/DP IV in-
formin (approx. 3 mol/l) appeared to moderately re-
teractions. We have been unable to support the claim
verse the loss in detection to 12% and 55% respectively.
that metformin inhibits DP IV activity by a number of
These findings compelled us to perform in vitro exper-
different experimental approaches. The most likely ex-
iments using alternative enzymological methods to re-
planation for the findings of Mannucci et al. (16) with
assess the role of metformin on DP IV.
respect to preservation of N-terminally intact GLP-1
In contrast to the in vitro findings of Mannucci et al.
with metformin treatment is that metformin increases
(16), we were unable to detect any significant effect of
the secretion of total GLP-1, and thus a proportional
metformin on Gly-Pro-pNA hydrolysis, the prototypical
increase in intact GLP-1 would be expected. It is diffi-
DP IV substrate, in healthy human serum, purified pig
cult to explain the disparate findings in vitro between
kidney DP IV or recombinant human DP IV. Further
the current report and that published previously, how-
experiments using MALDI-TOF mass spectrometry
ever, the primary experimental omission of measuring
which can concurrently detect disappearance and ap-
pearance of the molecular species corresponding to
mines the earlier data, as sample recovery cannot be
assessed. In contrast, MALDI-TOF mass spectrometry
allowed direct detection of both intact and inactive
exponential decay curves which can be compared under
GLP-1, and hence is more convincing. Surface plasmon
different experimental conditions. Consistent with
resonance allowed measurement of affinity of interac-
Gly-Pro-pNA enzymological experiments, metformin
tion between enzyme and substrate. This was not sig-
did not alter the degradation kinetics of GLP-1
nificantly altered by metformin or the presence of an
over a wide range of concentrations in any of the en-
In conclusion, it appears that metformin may in-
Surface plasmon resonance (SPR) was used to exam-
crease hormone secretion from both the pancreatic al-
ine the interaction between purified DP IV homologs
pha cell and intestinal L-cell, resulting in greater glu-
and GLP-1, irrespective of catalytic activity. The
cagon and total GLP-1 levels in metformin treated
amine-coupling reaction does not affect enzyme activ-
individuals. The latter effect may be one of the mech-
ity (19), and SPR allows measurement of intermolecu-
anisms by which metformin improves glucose toler-
lar interactions not necessarily confined to the cata-
ance. With the emergence of potent specific DP IV
lytic site. From these studies, apparent K values were
inhibitors for the treatment of type 2 diabetes mellitus,
an even greater potential may lie in combinatorial
formin failed to alter the binding interaction between
treatment with both metformin and DP IV inhibitors to
Binding constants for N-terminally truncated glucagonfragments and purified DP IV were not previously
This work was funded in part by Department of Science and
tested, however, studies indicated that substitution or
Technology of Sachsen Anhalt (HUD Grant 9704/00116) and by the
modification of the penultimate amino acid of glucagon
Medical Research Council of Canada (CHSM and RAP Grant590007) and the Canadian Diabetes Association. Simon Hinke is
reduced the apparent K by approximately 10-fold, and
grateful for the support of the Killam Trusts, the Medical Research
that altering the chirality of Gln3 produced more pro-
Council of Canada, and the Deutscher Akademischer Austausch-
nounced effects on glucagon/DP IV interactions (19).
dienst (DAAD). The authors thank Madeleine Speck, Joachim
Studies comparing DP IV binding constants for GIP
Ba¨r, Michael Wermann, and Dr. Susanne Manhart for technical
ϭ 1.7 mol/l) and DP IV hydrolysis product, assistance.
(K ϭ 3.2 mol/l), resulted in similar findings
1. (1999) Diabetes Statistics. NIH Publication No. 99-3892.
shown are one order of magnitude greater than
2. DeFronzo, R. A. (1999) Pharmacologic therapy for type 2 diabe-
those for GIP, measured under identical conditions.
tes mellitus. Ann. Intern. Med. 131, 281–303.
However, similar to GIP, N-terminal truncation does
3. Zhang, B. B., and Moller, D. E. (2000) New approaches in the
not dramatically affect binding affinity for DP IV.
treatment of type 2 diabetes. Curr. Opin. Chem. Biol. 4, 461–
Hence, it can be concluded that the substrate binding
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
4. Kieffer, T. J., McIntosh, C. H. S., and Pederson, R. A. (1995)
14. Bailey, C. J., and Turner, R. C. (1996) Metformin. N. Engl.
Degradation of glucose-dependent insulinotropic polypeptide
J. Med. 334, 574 –579.
and truncated glucagon-like peptide 1 in vitro and in vivo by
15. DeFronzo, R. A., and Goodman, A. M. (1995) Efficacy of met-
dipeptidyl peptidase IV. Endocrinology 136, 3585–3596.
formin in patients with non-insulin-dependent diabetes mellitus.
5. Unger, R. H., and Eisentraut, A. M. (1969) The entero-insular
N. Engl. J. Med. 333, 541–549.
axis. Arch. Intern. Med. 123, 261–266.
16. Mannucci, E., Ognibene, A., Cremasco, F., Bardini, G., Mencucci,
6. D’Alessio, D. (1997) Peptide hormone regulation of islet cell.
A., Pierazzuoli, E., Ciani, S., Messeri, G., and Rotella, C. M. Horm. Metab. Res. 29, 297–300.
(2001) Effect of metformin on glucagon-like peptide 1 (GLP-1)
7. Mentlein, R., Gallwitz, B., and Schmidt, W. E. (1993) Dipeptidyl-
and leptin levels in obese nondiabetic subjects. Diabetes Care 24,
peptidase IV hydrolyses gastric inhibitory polypeptide, glucagon-
like peptide-1 (7-36)amide, peptide histidine methionine and is
17. Mannucci, E., Ognibene, A., Cremasco, F., Bardini, G., Mencucci,
responsible for their degradation in human serum. Eur. J. Bio-
A., Pierazzuoli, E., Ciani, S., Messeri, G., and Rotella, C. M. chem. 214, 829 – 835.
(2000) Effects of metformin on baseline and oral glucose-induced
8. Pauly, R. P., Rosche, F., Wermann, M., McIntosh, C. H. S.,
glucagon-like peptide-1 (GLP-1) in obese non-diabetic subjects,
Pederson, R. A., and Demuth, H.-U. (1996) Investigation of
Abstract #472. Diabetes 49 (Suppl. 1).
GIP1– 42 and GLP-1 7-36 degradation in vitro by dipeptidylpeptidase IV (DPIV) using Matrix-Assisted Laser Desorption/
18. Wolf, B., Fischer, G., and Barth, A. (1978) [Kinetics of dipeptidyl-
Ionization—Time of Flight Mass Spectrometry (MALDI-TOF
peptidase IV] (German). Acta Biol. Med. Ger. 37, 409 – 420.
MS): A novel kinetic approach. J. Biol. Chem. 271, 23222–23229.
19. Hinke, S. A., Pospisilik, J. A., Demuth, H.-U., Mannhart, S.,
9. Pederson, R. A., White, H. A., Schlenzig, D., Pauly, R. P., McIn-
¨ hn-Wache, K., Hoffmann, T., Nishimura, E., Pederson, R. A.,
tosh, C. H. S., and Demuth, H.-U. (1998) Improved glucose tol-
and McIntosh, C. H. S. (2000) Dipeptidyl peptidase IV degrada-
erance in zucker fatty rats by oral administration of the dipep-
tion of glucagon: Characterization of glucagon degradation prod-
tidyl peptidase IV inhibitor isoleucine thiazolidide. Diabetes 47,
ucts and DPIV resistant analogs. J. Biol. Chem. 275, 3827–3834.
20. Rosche, F., Schmidt, J., Hoffmann, T., Pauly, R. P., McIntosh,
10. Pauly, R. P., Demuth, H.-U., Rosche, F., Schmidt, J., White,
C. H. S., Pederson, R. A., and Demuth, H. U. (2000) Kinetic
H. A., Lynn, F., McIntosh, C. H. S., and Pederson, R. A. (1999)
analysis of enzymatic and nonenzymatic degradation of peptides
Improved glucose tolerance in rats treated with the dipeptidyl
by MALDI-TOF MS. Methods Mol. Biol. 146, 251–272.
peptidase IV (CD26) inhibitor ile-thiazolidide. Metabolism 48,
21. Scheen, A. J. (1996) Clinical pharmacokinetics of metformin. Clin. Pharmacokinet. 30, 359 –371.
11. Ahren, B., Holst, J. J., Martensson, H., and Balkan, B. (2000)
22. Lugari, R., Dell’Anna, C., Sarti, L., Coppi, S., Verlato, C. A.,
Improved glucose tolerance and insulin secretion by inhibition of
Sbordone, P., Bianco, M., Gnudi, A., and Zandomeneghi, R.
dipeptidyl peptidase IV in mice. Eur. J. Pharmacol. 404, 239 –
(1998) Effects of metformin on intestinal and pancreatic endo-
crine secretion in type 2 (non-insulin-dependent) diabetes. In
12. Deacon, C. F., Hughes, T. E., and Holst, J. J. (1998) Dipeptidyl
Molecular and Cell Biology of Type 2 Diabetes and Its Compli-
peptidase IV inhibition potentiates the insulinotropic effect of
cations (Belfiore, F., Lorenzi, M., Molinatti, G. M., and Porta, M.,
glucagon-like peptides 1 in the anesthetized pig. Diabetes 47,
Eds.), pp. 161–163, Karger, Basel.
13. Balkan, B., Kwasnik, L., Miserendino, R., Holst, J. J., and Li, X.
23. Molloy, A. M., Ardill, J., and Tomkin, G. H. (1980) The effect of
(1999) Inhibition of dipeptidyl peptidase IV with NVP-DPP728
metformin treatment on gastric acid secretion and gastrointes-
increases plasma GLP-1 (7-36 amide) concentrations and im-
tinal hormone levels in normal subjects. Diabetologia 19, 93–96.
proves oral glucose tolerance in obese Zucker rats. Diabetologia
24. Kieffer, T. J., and Habener, J. F. (1999) The glucagon-like pep-
42, 1324 –1331.
tides. Endocr. Rev. 20, 876 –913.
Recent Developments in DNA-Computing Am Exerzierplatz 3, 97072 W¨urzburg, Germany Abstract computing time to solve the task are in good proportion. Adleman’s procedure uses polynomial computing time, but In 1994 Adleman published the description of a lab ex- so far there are only exponential time algorithms known to periment, where he computed an instance of the Hamilto- compute the H
Jahresbericht wird mit einer grosser Mehrheit angenommen. „nichts haltet mich zurück und was ist so schlimm mit ein paar Mordfällen vor Rechnung 2005 dem Z’vieri“ Blick so leise wie eine Treslag: Startgelderhöhung, Karten etwas günstiger + Wirtschaft ASJM-Schlusslauf kommt ins neue Vereinsjahr Sport-Toto Kartenbeitrag Buechberg erst 2006 Kartenverkauf Zurzacher kommt auch

 Biochemical and Biophysical Research Communications 291, 1302–1308 (2002)
Biochemical and Biophysical Research Communications 291, 1302–1308 (2002)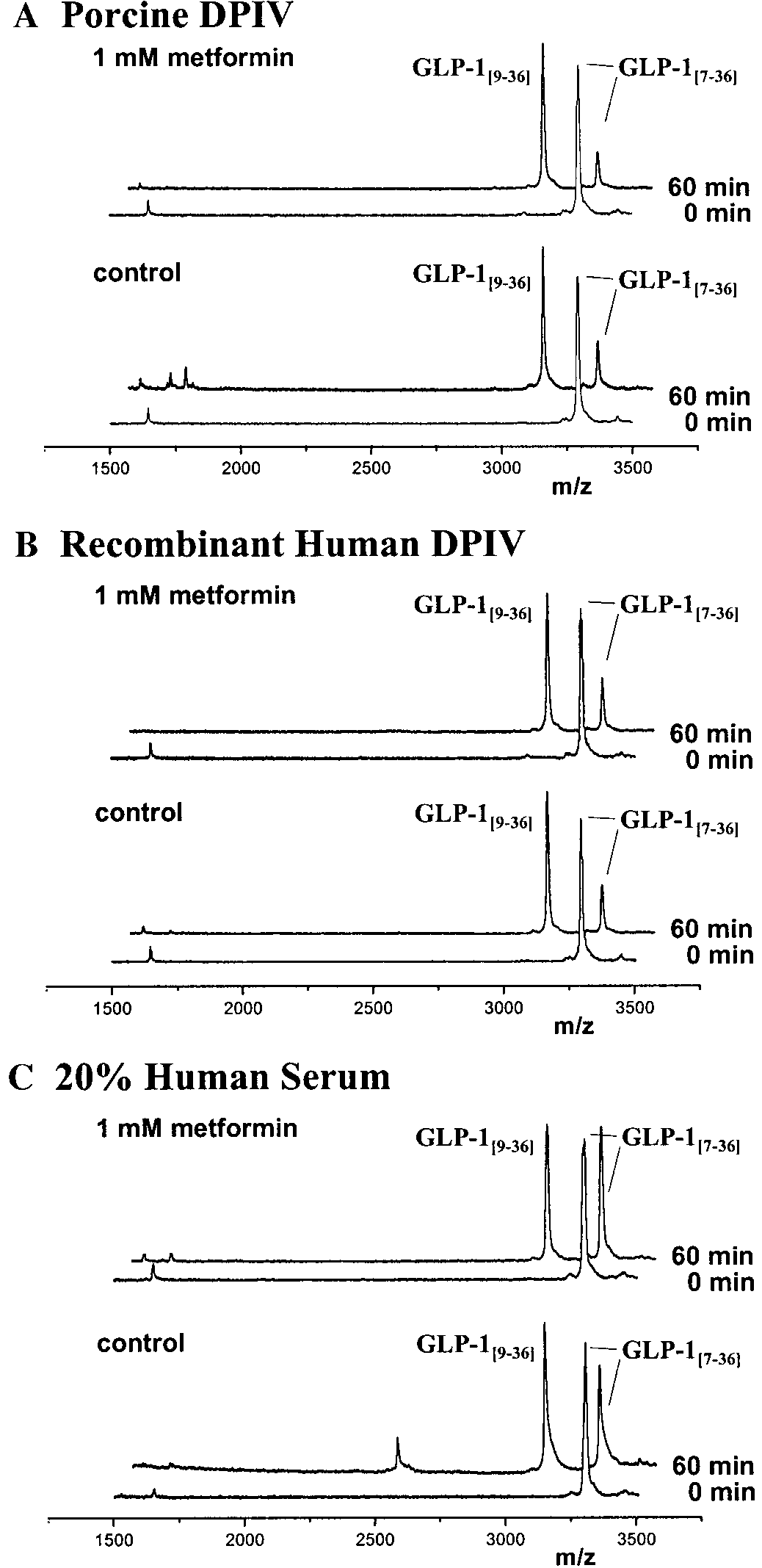 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Metformin fails to alter interaction between GLP-
Spectrophotometric Studies Using Gly-Pro-pNA
surface plasmon resonance examined the apparent
Note. Hydrolysis was monitored under standard conditions as de-
scribed under Materials and Methods.
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Metformin fails to alter interaction between GLP-
Spectrophotometric Studies Using Gly-Pro-pNA
surface plasmon resonance examined the apparent
Note. Hydrolysis was monitored under standard conditions as de-
scribed under Materials and Methods.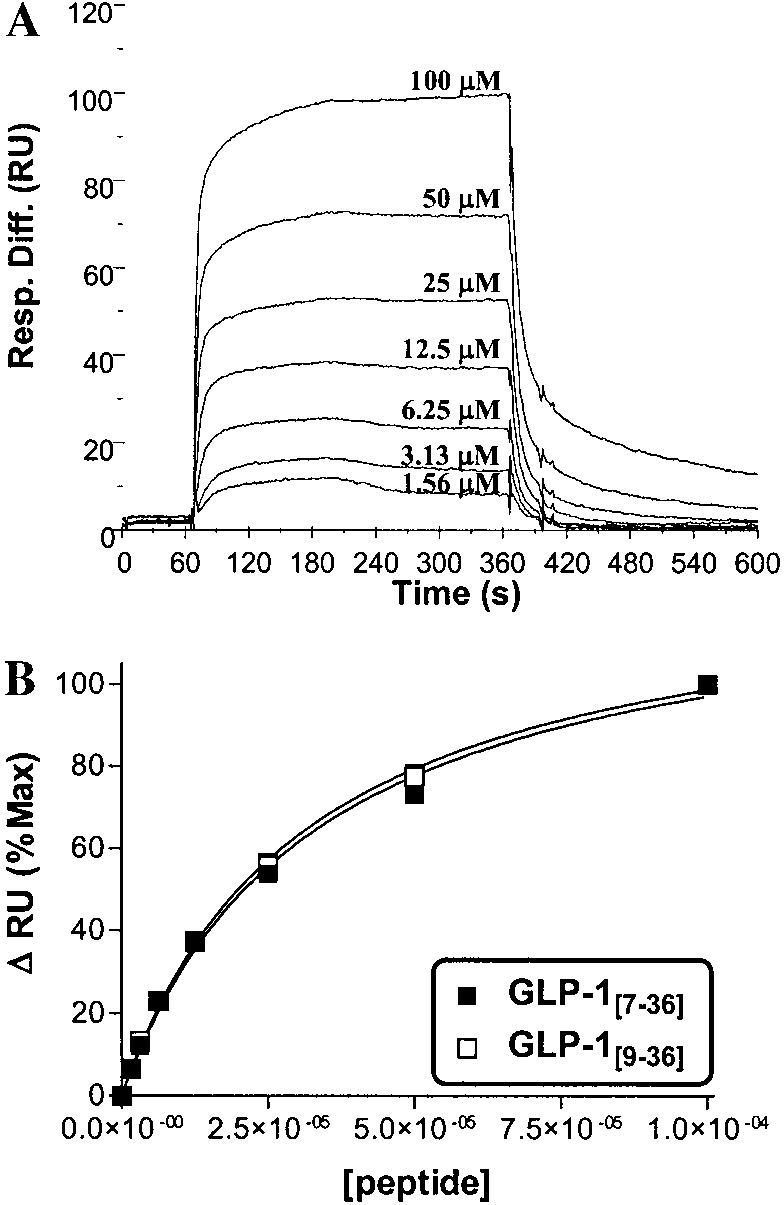 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Note. See text for detailed methods.
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Note. See text for detailed methods.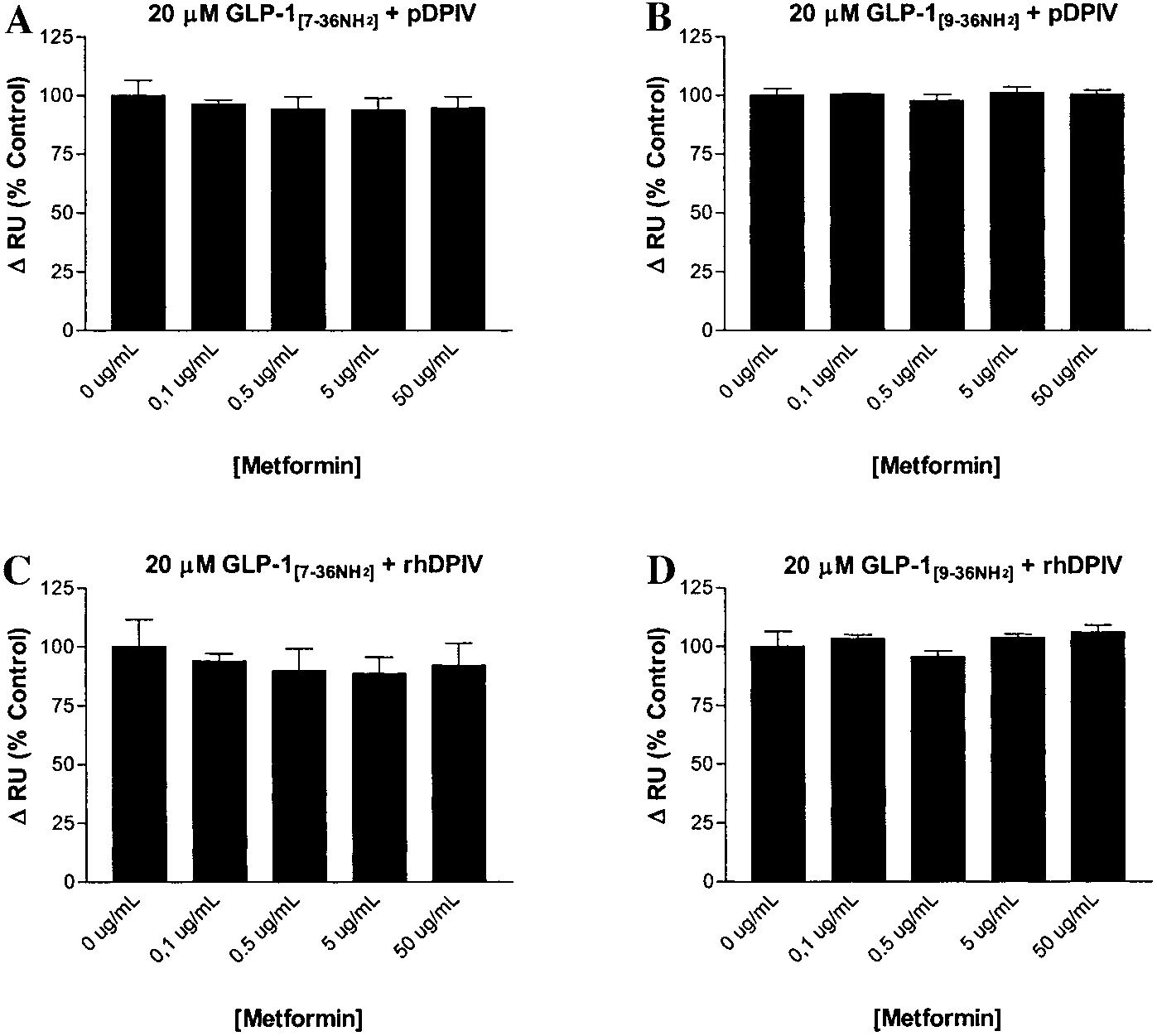 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Effect of graded metformin concentrations on interactions of GLP-1[7-36NH2] or GLP-1[9-36NH2] with porcine or recombinant human DP
similar experiments have not yet been carried out in
glucose sensitivity of the pancreatic alpha cell and
healthy or diabetic subjects, animal models, or in vitro.
enteroendocrine L-cell, or the secretory rate of these
We have addressed the latter deficiency, and per-
cells, resulting in greater hormone release with met-
formed several direct in vitro enzymological experi-
formin treatment. The work of Lugari et al. is sup-
ments to determine if metformin inhibits DP IV or
ported by studies published previously (23), which
alters the substrate-enzyme interaction. In contrast to
found that metformin significantly increased release of
findings by Mannucci and co-workers, we have been
pancreatic and gut glucagon (glicentin and oxynto-
unable to show that metformin has any effect on DP IV,
modulin intestinal products of proglucagon processing
and thus we offer alternative explanations for their
released in equal amounts to GLP-1 from enteroendo-
crine L-cells in response to luminal nutrients (24)).
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Effect of graded metformin concentrations on interactions of GLP-1[7-36NH2] or GLP-1[9-36NH2] with porcine or recombinant human DP
similar experiments have not yet been carried out in
glucose sensitivity of the pancreatic alpha cell and
healthy or diabetic subjects, animal models, or in vitro.
enteroendocrine L-cell, or the secretory rate of these
We have addressed the latter deficiency, and per-
cells, resulting in greater hormone release with met-
formed several direct in vitro enzymological experi-
formin treatment. The work of Lugari et al. is sup-
ments to determine if metformin inhibits DP IV or
ported by studies published previously (23), which
alters the substrate-enzyme interaction. In contrast to
found that metformin significantly increased release of
findings by Mannucci and co-workers, we have been
pancreatic and gut glucagon (glicentin and oxynto-
unable to show that metformin has any effect on DP IV,
modulin intestinal products of proglucagon processing
and thus we offer alternative explanations for their
released in equal amounts to GLP-1 from enteroendo-
crine L-cells in response to luminal nutrients (24)).