Kamagra enthält Sildenafilcitrat als pharmakologisch aktiven Bestandteil. Dieser hemmt selektiv die Phosphodiesterase-5 und erhöht dadurch die Konzentration von cGMP im Corpus cavernosum. Der Effekt ist zeitlich begrenzt, da die Halbwertszeit von Sildenafil etwa vier Stunden beträgt. In der galenischen Form als Mundgel erfolgt die Resorption besonders rasch, was zu einem schnelleren Wirkeintritt führt. Der Abbau erfolgt überwiegend hepatisch über CYP3A4, wobei ein aktiver Metabolit entsteht, der zur Gesamtwirkung beiträgt. Typische Nebenwirkungen ergeben sich aus der Vasodilatation, darunter leichte Kopfschmerzen und nasale Kongestion. In klinischen Beschreibungen wird kamagra oral jelly im Zusammenhang mit der schnelleren Absorption erwähnt.
Files.petparasitologiaufpe.webnode.com.br
INFECTION AND IMMUNITY, Feb. 2005, p. 1265–1269
0019-9567/05/$08.00ϩ0 doi:10.1128/IAI.73.2.1265–1269.2005Copyright 2005, American Society for Microbiology. All Rights Reserved. Lactobacillus johnsonii La1 Antagonizes Giardia intestinalis In Vivo
Martı´n A. Humen,1 Graciela L. De Antoni,1,2 Jalil Benyacoub,3 Marı´a E. Costas,4
Marta I. Cardozo,4 Leonora Kozubsky,4 Kim-Yen Saudan,3
´n y Desarrollo en Criotecnologı´a de Alimentos,1 and Ca´tedra de Microbiologı´a2 and CaParasitologı´a,4 Departamento de Ciencias Biolo´gicas, Facultad de Ciencias Exactas, Universidad Nacional deLa Plata, La Plata, Argentina, and Nestle´ Research Center, Vers-Chez-les-Blanc,
Received 23 August 2004/Returned for modification 16 September 2004/Accepted 18 October 2004
This study describes the in vivo activity of Lactobacillus johnsonii La1 (NCC533) in Giardia intestinalis- infected gerbils (Meriones unguiculatus). Daily administration of lactobacilli in the drinking water from 7 days before inoculation with Giardia trophozoites efficiently prevented G. intestinalis strain WB clone C6 from infecting gerbils. More specifically, shedding of fecal Giardia antigens (GSA65 protein) was diminished in the La1-treated group, and resolution of infection was observed by 21 days postinoculation. Histology and analysis of enzymatic markers of microvillus membrane integrity revealed that probiotic administration also protected against parasite-induced mucosal damage. In addition, a cellular response to Giardia antigens was stimulated in spleen cells from La1-treated gerbils. Results show for the first time the antigiardial effect of probiotic lactobacilli in vivo and provide further insight into the antagonistic properties of lactic acid bacteria against protozoa involved in intestinal infections. Giardia intestinalis, a flagellated protozoan, is the etiological
vivo model of giardiasis (Meriones unguiculatus) to gain further
agent of giardiasis, an intestinal infection which causes diar-
insight into the antigiardial properties of L. johnsonii La1
rhea and malabsorption in humans and animals (19, 22). This
(NCC533; Nestec Culture Collection). The protocol was ap-
parasite has a biphasic developmental cycle with two morpho-
´ Research Center and state ethical com-
logically different forms: cysts and trophozoites. The latter is
the vegetative form of the parasite which is responsible for
Frozen suspensions of L. johnsonii (hereafter La1) were
infection and pathological manifestations (19). Infection fol-
thawed and reactivated in MRS broth before the study. The
lows the ingestion of cysts which then give rise to trophozoites
bacteria were then subcultured in MRS broth for 16 h at 37°C
after passage through the stomach and the upper intestinal
and harvested by centrifugation at 10,000 ϫ g for 10 min.
Concentrated suspensions were frozen at Ϫ80°C with 10%
Giardia is generally treated with antibiotics (e.g., nitroimid-
(vol/vol) glycerol until required. After thawing, bacterial
azoles and nitrofurans). However, clinical failures, occurrence
counts were determined by plating serial dilutions on MRS
of resistant strains, and side effects of anti-Giardia drugs (21,
26, 30, 44) have encouraged research on alternative therapeu-
Trophozoites of clone C6 of the strain WB of G. intestinalis,
tic strategies which have included the use of plant extracts and
products derived from bees (12, 21, 25). However, since pro-
doba, Argentina, were grown in Keister’s modified TYI-S-33
biotic microorganisms provide health benefits to the host by
medium (28) as previously described (36) and then frozen in
antagonizing pathogens and modulating both innate and ac-
quired immunity at local and systemic levels (13, 23, 24, 33, 40),
Suspensions of La1 were prepared daily by diluting thawed
we wondered if such organisms could also be used in the
concentrated suspensions in mineral water. Young adult Mon-
treatment of parasitic infections. Certainly, colonization of the
golian gerbils (Meriones unguiculatus) ingested bacterial sus-
intestine by Giardia strongly depends on the intestinal micro-
pensions ad libitum in their drinking water each day beginning
flora (43). Furthermore, in vitro studies show that some pro-
7 days prior to inoculation with Giardia trophozoites. Based on
biotic lactobacilli (e.g., Lactobacillus johnsonii La1) are able to
the daily water intake and the bacterial concentration in drink-
interfere with the cell cycle of G. intestinalis (36). However,
ing water, the daily bacterial intake was estimated to be 108
since this antigiardial effect does not seem to be a general
CFU per animal. Control animals received mineral water in-
property of lactobacilli, further research is required to select
stead of the bacterial suspensions throughout the course of the
probiotic strains which can be used in the prevention and
study. Seven days after probiotic treatment had begun, frozen
treatment of such an infection. For this reason, we used an in
trophozoites were thawed in a water bath at 37°C, and a vol-ume of 200 l (5 ϫ 105 trophozoites) was administered bygavage to each animal in the two treatment groups. At various
* Corresponding author. Mailing address: CIDCA, Universidad Na-
time points thereafter, animals were killed by cervical disloca-
cional de La Plata, Calle 47 y 116 (CC 553), 1900 La Plata, Argentina.
tion, and the duodenal contents were analyzed for the presence
Phone: 54 221 424 92 87. Fax: 54 221 425 48 53. E-mail: pfp@nahuel.biol.unlp.edu.ar.
of trophozoites. To this end, 5-cm-long segments of the small
intestine were excised at a distance of 11 cm from the pylorus.
TABLE 1. Infection of gerbils at different time points after
These samples were then placed in 2 ml of ice-cold culture
inoculation with G. intestinalis WB clone C6a
medium, opened longitudinally, and vortexed. Enumeration of
trophozoites was done by using a hemocytometer with a de-
tection limit of 800 trophozoites/cm.
In order to determine fecal giardial antigens, five fecal pel-
lets (0.27 [Ϯ0.05] g [wet weight]) per gerbil were homogenized
in 0.5 ml of extraction buffer (50 mM EDTA, 100 g of soy-
bean trypsin inhibitor [Sigma, St. Louis, Mo.]/ml in phos-
phate-buffered saline), vortexed for 2 min, and centrifuged at
Gerbils received either La1 or placebo daily from 7 days before inoculation
with the trophozoites of G. intestinalis WB clone C6 (5 ϫ 105 trophozoites per
13,000 ϫ g for 15 min. Detection of GSA65 antigen in the
animal). Results are from four independent experiments.
supernatants (1) was performed by using a ProSpecT kit ac-
b Number of gerbils with detectable parasites in the small intestine/total num-
cording to the instructions of the manufacturer (Alexon-
ber of gerbils studied. The statistical significance of the infection rate of La1-treated animals compared to that of controls was calculated using Fisher’s exact
Trend). Cyst determination was performed microscopically on
test. P values are shown in parentheses. c Averages Ϯ standard deviations of the log of parasites per infected gerbil.
ND, no parasites were detected (Ͻ800 trophozoites/cm).
For histological analysis, 1 cm of the small intestine collected
10 cm from the pylorus was fixed in Bouin fixative and dehy-drated in a graded series of ethanol. Paraffin-embedded sec-tions of 5 m were rehydrated and stained with hematoxylin
pare infection rates, and the two-tailed t test with equal vari-
ances was used to compare mucosal sucrase activity and cell
Intestinal sucrase activity was measured in 5-cm-long sec-
proliferation indices of the two groups (Software InfoStat,
tions of the small intestine which were collected 5 cm below the
2004; Grupo InfoStat, FCA, Universidad Nacional de Co
pylorus. The intestinal samples were placed in ice-cold phos-
phate-buffered saline and flushed with 50 ml of ice-cold dis-
As shown in Table 1, there was a peak in the Giardia infec-
tilled water using a syringe. The segments were then opened
tion rate 7 days postinoculation. At this time point, 10 of the 12
longitudinally, and the mucosa was collected by scraping with
gerbils (83%) in the placebo group and 3 out of 11 gerbils in
a glass microscope slide. Scrapings were stored at Ϫ20°C until
the La1-treated group had trophozoites in their small intestine
analysis. Intestinal disaccharidase activity was determined as
(P ϭ 0.01). By day 14, the infection was resolved in the La1-
previously reported (16). Protein concentrations were deter-
treated group, but 6 out of 14 gerbils in the placebo group were
mined using the Bradford assay, and the results are expressed
still infected (Table 1) (P ϭ 0.02). Of these gerbils, three were
as arbitrary units per gram of protein (3).
still infected 21 days after inoculation. Although probiotic
To determine cell proliferation indices, spleens were placed
treatment reduced the infection rate, it had no significant effect
in 2 ml of complete RPMI 1640 medium (10% fetal calf serum,
on the intensity of infection (Table 1).
1% nonessential amino acids), homogenized through a cell
Fecal shedding of GSA65 antigen followed different kinetics
strainer (70 m; BD Falcon, Bedford, Mass.), and centrifuged
from that of live trophozoites in the small intestine. In the
at 250 ϫ g for 5 min. The pellets were then rapidly lysed with
placebo group, the number of GSA65-positive gerbils progres-
1 ml of sterile distilled water and centrifuged at 250 ϫ g for 5
sively increased from day 7 to day 21 post-Giardia inoculation
min. Cells were resuspended in complete RPMI 1640 medium
(Fig. 1). In contrast, the single gerbil positive for GSA65 an-
at a concentration of 106 cells/ml before testing cell prolifera-
tigen in the La1-treated group at day 14 was no longer positive
tion in the presence of the T-cell mitogen concanavalin A
(ConA) at a concentration of 3 g/ml or the B-cell mitogen
G. intestinalis infection in gerbils has a profound effect on
pokeweed mitogen (PWM) at a concentration of 1 g/ml (both
the structure and function of the intestinal mucosa (Fig. 2).
from Sigma). Stimulation in the presence of Giardia antigens
Villus shortening and hypercellularity of the lamina propria
was also evaluated. These membrane antigens were extracted
due to increased numbers of mononuclear, polymorphonu-
as reported previously (17) and used at a concentration of 10
clear, and eosinophilic cells were observed in control animals.
In contrast, no morphological damage of the gut mucosa was
Spleen cells (105 cells/well) were incubated with complete
observed in La1-treated animals. These findings correlated
RPMI 1640 medium (control) supplemented with either
well with the difference in mucosal sucrase activity observed in
ConA, PWM, or Giardia antigen in a final volume of 200 l of
the two groups. While the intestines of La1-treated animals
medium. After incubation at 37°C for 72 h, the cells were pulse
maintained an enzymatic activity of approximately 200 arbi-
labeled with 1.5 Ci of [3H]thymidine (Amersham Pharmacia
trary units/g of protein throughout the study, the intestines of
¨bendorf, Switzerland) for a further 16 h. The cells
control animals had decreasing enzymatic activity over time
were then harvested on nitrocellulose filters (Packard Bio-
(Fig. 3). Differences between the two groups were statistically
sciences B.V., Groningen, The Netherlands), and the incorpo-
significant at both 14 and 21 days postinoculation (P Ͻ 0.05).
ration of [3H]thymidine was measured by scintillation counting
No differences in spleen cell proliferation were observed
(TopCount; Packard Biosciences B.V.). Stimulation indices
between the two treatment groups 7 days after inoculation with
(SI) were determined as the severalfold increase in titrated
Giardia trophozoites (data not shown). However, at day 14,
thymidine incorporation in stimulated cells compared to that
splenocytes of La1-treated animals showed a greater response
to ConA (placebo group SI, 7.8 Ϯ 2.2; La1-treated group SI,
Fisher’s two-tailed exact statistical analysis was used to com-
22.8 Ϯ 11.6 [P ϭ 0.09]), to PWM, and to Giardia antigen
FIG. 3. Intestinal sucrase activity. Gerbils received either La1 or
FIG. 1. Shedding of fecal GSA65 antigen. Gerbils received either
placebo daily from 7 days before inoculation with the trophozoites of
La1 (■) or placebo (E) daily from 7 days before inoculation with the
G. intestinalis WB clone C6 (5 ϫ 105 trophozoites per animal). Values
trophozoites of G. intestinalis WB clone C6 (5 ϫ 105 trophozoites per
(arbitrary units [AU] per gram of protein) represent averages from six
animal). Values represent the ratio between GSA65-positive (GSA65
gerbils. Error bars indicate standard deviations. Differences between
ϩ) gerbils and the total number of gerbils at various times after
La1 and placebo groups are statistically significant (P ϭ 0.05 and P ϭ
Giardia infection. The number of animals per group was 18, 12, and 5
0.009 at 14 and 21 days, respectively).
at 7, 14, and 21 days post-Giardia infection, respectively.
Ϯ 0.9 [P ϭ 0.007]) and to the B-cell mitogen PWM (placebo
(placebo group SI, 1.2 Ϯ 0.2; La1-treated group SI, 3.7 Ϯ 0.8
group SI, 2.1 Ϯ 0.6; La1-treated group SI, 3.6 Ϯ 0.6 [P ϭ 0.03])
[P ϭ 0.006]) than those of control animals. At 21 days post-
was observed, but there was no longer a difference in the
Giardia infection, a significantly greater response to Giardia
cellular response to ConA between the two groups (data not
antigen (placebo group SI, 0.9 Ϯ 0.2; La1-treated group SI, 3.6
shown). Research on the efficacy of probiotic bacteria against
FIG. 2. Histological sections of small intestine. Gerbils received either La1 (A) or placebo (B) daily from 7 days before inoculation with the
trophozoites of G. intestinalis WB clone C6 (5 ϫ 105 trophozoites per animal). Intestinal segments were collected 7 days post-Giardia infection. Sections were stained with hematoxylin and eosin and observed at ϫ100 original magnification. (C) Further magnification of the intestine froma control animal. Arrows indicate the presence of trophozoites.
intestinal infections has focused primarily on protection
different variant surface proteins in control and La1-treated
against bacterial pathogens. The present in vivo study consol-
groups could not be ruled out (35), our results suggest a prim-
idates recent reports describing the capacity of some lactoba-
ing of the immune system by La1 that leads to clearance of
cilli of human and canine origin to antagonize Giardia infec-
both trophozoites and Giardia antigens from the intestines
tion which interferes with the growth and cell cycle of G.
(Table 1 and Fig. 1). It is possible that this reflects the devel-
intestinalis in vitro (36). Mongolian gerbils (Meriones unguicu-
opment of a memory response following probiotic administra-
latus) are a good animal model for studying Giardia infection
with strains of human origin (3, 4, 11). Their susceptibility to
Although the mechanisms underlying the virulence of G.
giardiasis has been attributed to a deficient antibody response
intestinalis remain poorly understood, both direct interaction
to specific Giardia antigens (34). We have observed that gerbils
with enterocytes and secretion of soluble mediators are
are readily infected with trophozoites administered by gavage
thought to contribute to the pathogenesis (14). The immune
and begin to eliminate cysts in their feces 1 week after inocu-
response of the host and inflammatory status of the intestinal
lation (data not shown). As a consequence of this infection, the
mucosa are also key determinants (41). It is therefore inter-
animals have reduced disaccharidase activity in their intestinal
esting that a lower cellular infiltration was observed in the
intestinal villi of La1-treated animals than that of controls (Fig.
The presence of viable trophozoites in the small intestine is
2). This result indicates the absence of inflammatory processes
a recognized marker of Giardia infection (37, 42). We found
in the intestine of La1-treated gerbils.
that administration of L. johnsonii La1 dramatically reduces
In the present study, it is unlikely that metabolic products in
the proportion of gerbils with active trophozoites in the gut
the bacterial suspensions contribute to the protective effects
and leads to resolution of infection within 14 days. In contrast,
observed. Although lactic acid is the main product of La1
viable trophozoites were consistently observed in the intestines
metabolism, only traces of this organic acid were detected in
of the placebo group at all the time points studied (Table 1).
the bacterial suspensions administered (data not shown). Thus,
Furthermore, we have found a time-dependent increase in the
the antigiardial effect of L. johnsonii La1 in vivo is probably
proportion of GSA65-positive gerbils in this group (Fig. 1).
due to the production of inhibitory substances in situ and/or to
Since GSA65 antigen is present on both trophozoites and cysts
the modulation of the immune response of the host. Certainly,
(38), it is important that the increased proportion of GSA65-
previously published results have shown that L. johnsonii La1
positive gerbils with time may reflect changes in the numbers
mediates its anti-Giardia effect through the production of sol-
of both trophozoites and cysts reaching the distal small intes-
uble mediators (36) and that it is able to modulate the immune
tine. Certainly, differences in the kinetics of both GSA65 shed-
ding and trophozoite load in the small intestine are related to
To summarize, we report for the first time an in vivo effect
the distribution of trophozoites and cysts along the orad-
of a probiotic lactobacillus against G. intestinalis. Our results
provide scientific evidence that the applications of probiotic
Progression of Giardia infection in gerbils correlates with a
organisms can be extended to include alternative strategies in
progressive impairment in intestinal sucrase activity (3), which
the prevention of a widespread parasitic infection.
may account for the diminished nutrient absorption and failureto thrive associated with giardiasis (11, 19). However, admin-
We are indebted to the Agencia de Promocio
istration of L. johnsonii La1 protects against the impaired
´gica for financial support. M.A.H. is a fellow at the Consejo Nacional
sucrase activity and the histological changes found in infected
member of the Carrera del Investigador Cientı´fico y Tecnolo
animals. (Fig. 2). It is noteworthy that cysts were eliminated in
´n de Investigaciones Cientı´ficas de la Provincia de Buenos
only a few of the gerbils in the placebo group (data not shown).
Aires (CIC-PBA), Buenos Aires, Argentina; and P.F.P. is a member of
This is an important observation because cyst production is the
the Carrera del Investigador Cientı´fico y Tecnolo
only means by which the parasite is known to spread. This
observation is also in agreement with previous studies in vitro
C6. We also thank Martine Rouvet for her help in the preparation of
which have demonstrated that extracellular factors from La1
photographic images, Samuel Steffen and Lucı´a Brandy for technical
arrest G. intestinalis in the G phase of the cell cycle (36).
assistance, and Alicia N. Califano for statistical assessment. Revision
According to Bernander et al. (6), only trophozoites in G
of the manuscript by Anne Donnet is greatly acknowledged.
The protective effect of probiotics against intestinal patho-
REFERENCES
gens has been ascribed to many factors including antagonism
1. Addis, D. G. 1991. Evaluation of a commercially available enzyme-linked
by extracellular factors (8, 15, 31), interference with pathogen-
immunosorbent assay for Giardia lamblia antigen in stool. J. Clin. Microbiol. 29:1137–1142.
enterocyte interactions (7, 9), and modulation of the immune
2. Aley, S. B., M. Zimmerman, M. Hetsko, M. E. Selsted, and F. D. Gillin. 1994.
response (5, 23, 39). Not only innate defense mechanisms but
Killing of Giardia lamblia by cryptdins and cationic neutrophil peptides.
both acquired humoral and cell-mediated immune responses
Infect. Immun. 62:5397–5403.
3. Belosevic, M., G. M. Faubert, and J. D. MacLean. 1989. Disaccharidase
protect against Giardia infection (2, 18, 20, 27, 28, 42). Here,
activity in the small intestine of gerbils (Meriones unguiculatus) during pri-
we suggest that La1 treatment reinforces the immune response
mary and challenge infections with Giardia lamblia. Gut 30:1213–1219.
against Giardia in inoculated gerbils. More specifically, we
4. Belosevic, M., G. M. Faubert, J. D. MacLean, C. Law, and N. A. Croll. 1983. Giardia lamblia infections in Mongolian gerbils: an animal model. J. Infect.
show an increased splenocyte response to a T-cell mitogen and
Dis. 147:222–226.
to Giardia antigens at 14 days postinfection and an increased
5. Benyacoub, J., G. L. Maulden, C. Cavadini, T. Sauthier, R. E. Anderson, E. J. Schiffrin, and T. der Weid. 2003. Supplementation of food with En-
response to a B-cell mitogen and to Giardia antigens at 21 days
terococcus faecium (SF68) stimulates immune functions in young dogs. J.
postinfection. Even though selection of trophozoites bearing
Nutr. 133:1158–1162.
6. Bernander, R., J. E. Palm, and S. G. Svard. 2001. Genome ploidy in different
26. Katelaris, P. H., A. Naeem, and M. J. Farthing. 1994. Activity of metroni-
stages of the Giardia lamblia life cycle. Cell. Microbiol. 3:55–62.
dazole, azithromycin and three benzimidazoles on Giardia lamblia growth
7. Bernet, M. F., D. Brassart, J. R. Neeser, and A. L. Servin. 1994. Lactobacillus
and attachment to a human intestinal cell line. Aliment. Pharmacol. Ther. acidophilus La1 binds to cultured human intestinal cell lines and inhibits cell
8:187–192.
attachment and cell invasion by enterovirulent bacteria. Gut 35:483–489.
27. Kaur, H., H. Samra, S. Ghosh, V. K. Vinayak, and N. K. Ganguly. 1999.
8. Bernet-Camard, M. F., V. Lievin, D. Brassart, J. R. Neeser, A. L. Servin, and
Immune effector responses to an excretory-secretory product of GiardiaS. Hudault. 1997. The human Lactobacillus acidophilus strain La1 secretes a lamblia. FEMS Immunol. Med. Microbiol. 23:93–105.
nonbacteriocin antibacterial substance(s) active in vitro and in vivo. Appl.
28. Keister, D. B. 1983. Axenic culture of Giardia lamblia in TYI-S-33 medium
Environ. Microbiol. 63:2747–2753.
supplemented with bile. Trans. R. Soc. Trop. Med. Hyg. 77:487–488.
9. Bibiloni, R., P. F. Pe´rez, and G. L. De Antoni. 1999. Will a high adhering
29. Langford, T. D., M. P. Housley, M. Boes, J. Chen, M. F. Kagnoff, F. D. Gillin,
capacity in a probiotic strain guarantee exclusion of pathogens from intes-
and L. Eckmann. 2002. Central importance of immunoglobulin A in host
tinal epithelia? Anaerobe 5:519–524.
defense against Giardia spp. Infect. Immun. 70:11–18.
10. Blum, S., S. Alvarez, D. Haller, P. Perez, and E. J. Schiffrin. 1999. Intestinal
30. Lemee, V., I. Zaharia, G. Nevez, M. Rabodonirina, P. Brasseur, J. J. Ballet,
microflora and the interaction with immunocompetent cells. Antonie Leeu-
and L. Favennec. 2000. Metronidazole and albendazole susceptibility of 11
wenhoek 76:199–205.
clinical isolates of Giardia duodenalis from France. J. Antimicrob. Che-
11. Buret, A., J. A. Hardin, M. E. Olson, and D. G. Gall. 1992. Pathophysiology
mother. 46:819–821.
of small intestinal malabsorption in gerbils infected with Giardia lamblia.
31. Lievin-Le Moal, V., R. Amsellem, A. L. Servin, and M. H. Coconnier. 2002.
Gastroenterology 103:506–513. Lactobacillus acidophilus (strain LB) from the resident adult human gastro-
12. Calzada, F., C. M. Cerda-Garcia-Rojas, M. Meckes, R. Cedillo-Rivera, R.
intestinal microflora exerts activity against brush border damage promoted
Bye, and R. Mata. 1999. Geranins A and B, new antiprotozoal A-type
by a diarrhoeagenic Escherichia coli in human enterocyte-like cells. Gut
proanthocyanidins from Geranium niveum. J. Nat. Prod. 62:705–709. 50:803–811.
13. Cebra, J. J. 1999. Influences of microbiota on intestinal immune system
32. Link-Amster, H., F. Rochat, K. Y. Saudan, O. Mignot, and J. M. Aeschli-
development. Am. J. Clin. Nutr. 69:1046S–1051S. mann. 1994. Modulation of a specific humoral immune response and
14. Chin, A. C., D. A. Teoh, K. G.-E. Scott, J. B. Meddings, W. K. Macnaughton,
changes in intestinal flora mediated through fermented milk intake. FEMS
and A. G. Buret. 2002. Strain-dependent induction of enterocyte apoptosis by
Immunol. Med. Microbiol. 10:55–63. Giardia lamblia disrupts epithelial barrier function in a caspase-3-dependent
33. Miettinen, M., J. Vuopio-Varkila, and K. Varkila. 1996. Production of hu-
manner. Infect. Immun. 70:3673–3680.
man tumor necrosis factor alpha, interleukin-6, and interleukin-10 is induced
15. Coconnier, M. H., V. Lievin, E. Hemery, and A. L. Servin. 1998. Antagonistic
by lactic acid bacteria. Infect. Immun. 64:5403–5405.
activity against Helicobacter infection in vitro and in vivo by the human
34. Mohanty, M. C., and B. Ravindran. 2002. Deficiency of antibody responses Lactobacillus acidophilus strain LB. Appl. Environ. Microbiol. 64:4573–4580.
to T-independent antigens in gerbils—Meriones unguiculatus. Dev. Comp.
16. Dahlqvist, A. 1968. Assay of intestinal disaccharidases. Anal. Biochem. 22:
Immunol. 26:385–391.
35. Nash, T. E. 2002. Surface antigen variation in Giardia lamblia. Mol. Micro-
17. Dunn, L. A., J. A. Upcroft, E. V. Fowler, B. S. Matthews, and P. Upcroft.
biol. 45:585–590.
2001. Orally administered Giardia duodenalis extracts enhance an antigen-
36. Pe´rez, P. F., J. Minnaard, M. Rouvet, C. Knabenhans, D. Brassart, G. L. De
specific antibody response. Infect. Immun. 69:6503–6510. Antoni, and E. J. Schiffrin. 2001. Inhibition of Giardia intestinalis by extra-
18. Eckmann, L., F. Laurent, T. D. Langford, M. L. Hetsko, J. R. Smith, M. F.
cellular factors from lactobacilli: an in vitro study. Appl. Environ. Microbiol. Kagnoff, and F. D. Gillin. 2000. Nitric oxide production by human intestinal 67:5037–5042.
epithelial cells and competition for arginine as potential determinants of hostdefense against the lumen-dwelling pathogen Giardia lamblia. J. Immunol.
37. Roberts-Thomson, I. C., D. P. Stevens, A. A. Mahmoud, and K. S. Warren.
1976. Giardiasis in the mouse: an animal model. Gastroenterology 71:57–61. 164:1478–1487.
19. Farthing, M. J. G. 1995. Giardia lamblia, p. 1081–1105. In M. J. Blaser, P. D.
38. Rosoff, J. D., and H. H. Stibbs. 1986. Isolation and identification of a Giardia
Smith, J. I. Ravdin, H. B. Greenberg, and R. L. Guerrant (ed.), Infections of
lamblia-specific stool antigen (GSA 65) useful in coprodiagnosis of giardia-
the gastrointestinal tract. Raven Press, New York, N.Y.
sis. J. Clin. Microbiol. 23:905–910.
20. Faubert, G. 2000. Immune response to Giardia duodenalis. Clin. Microbiol.
39. Schiffrin, E. J., D. Brassart, A. L. Servin, F. Rochat, and A. Donnet-Hughes.
Rev. 13:35–54.
1997. Immune modulation of blood leukocytes in humans by lactic acid
21. Gardner, T. B., and D. R. Hill. 2001. Treatment of giardiasis. Clin. Microbiol.
bacteria: criteria for strain selection. Am. J. Clin. Nutr. 66:515S–520S.
Rev. 14:114–128.
40. Schiffrin, E. J., F. Rochat, H. Link-Amster, J. M. Aeschlimann, and A.
22. Gillin, F. D., D. S. Reiner, and J. M. McCaffery. 1996. Cell biology of the Donnet-Hughes. 1995. Immunomodulation of human blood cells following
primitive eukaryote Giardia lamblia. Annu. Rev. Microbiol. 50:679–705.
the ingestion of lactic acid bacteria. J. Dairy Sci. 78:491–497.
23. Haller, D., S. Blum, C. Bode, W. P. Hammes, and E. J. Schiffrin. 2000.
41. Scott, K. G., M. R. Logan, G. M. Klammer, D. A. Teoh, and A. G. Buret.
Activation of human peripheral blood mononuclear cells by nonpathogenic
2000. Jejunal brush border microvillous alterations in Giardia muris-infected
bacteria in vitro: evidence of NK cells as primary targets. Infect. Immun.
mice: role of T lymphocytes and interleukin-6. Infect. Immun. 68:3412–3418. 68:752–759.
42. Singer, S. M., and T. E. Nash. 2000. T-cell-dependent control of acute
24. Isolauri, E., Y. Sutas, P. Kankaanpaa, H. Arvilommi, and S. Salminen. 2001. Giardia lamblia infections in mice. Infect. Immun. 68:170–175.
Probiotics: effects on immunity. Am. J. Clin. Nutr. 73:444S–450S.
43. Singer, S. M., and T. E. Nash. 2000. The role of normal flora in Giardia
25. Johns, T., G. M. Faubert, J. O. Kokwaro, R. L. Mahunnah, and E. K. lamblia infections in mice. J. Infect. Dis. 181:1510–1512. Kimanani. 1995. Anti-giardial activity of gastrointestinal remedies of the
44. Upcroft, P., and J. A. Upcroft. 2001. Drug targets and mechanisms of resis-
Luo of east Africa. J. Ethnopharmacol. 46:17–23.
tance in the anaerobic protozoa. Clin. Microbiol. Rev. 14:150–164.
Heartburn Hiatus hernia is a common finding at gastroscopy AND MAY CAUSE NO SYMPTOMS. What Is HEARTBURN (gastro-oesophageal reflux)? Gastroesophageal reflux is the return of the stomach's contents back up into the oesophagus. There is a sphincter, or muscle in the lower oesophagus. Normally, the sphincter opens to allow food to pass into the stomach and closes to prevent ac
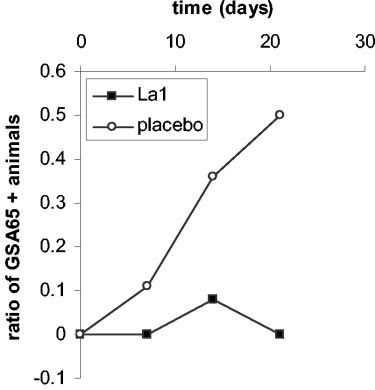
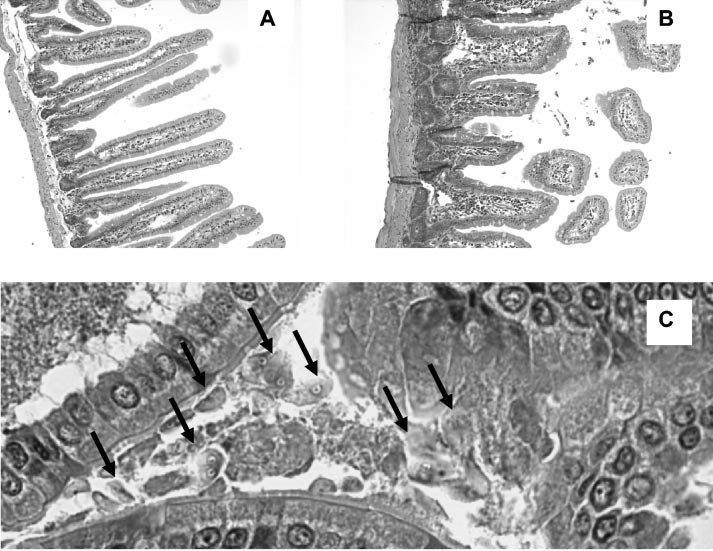
 FIG. 3. Intestinal sucrase activity. Gerbils received either La1 or
FIG. 1. Shedding of fecal GSA65 antigen. Gerbils received either
placebo daily from 7 days before inoculation with the trophozoites of
La1 (■) or placebo (E) daily from 7 days before inoculation with the
G. intestinalis WB clone C6 (5 ϫ 105 trophozoites per animal). Values
trophozoites of G. intestinalis WB clone C6 (5 ϫ 105 trophozoites per
(arbitrary units [AU] per gram of protein) represent averages from six
animal). Values represent the ratio between GSA65-positive (GSA65
gerbils. Error bars indicate standard deviations. Differences between
ϩ) gerbils and the total number of gerbils at various times after
La1 and placebo groups are statistically significant (P ϭ 0.05 and P ϭ
Giardia infection. The number of animals per group was 18, 12, and 5
0.009 at 14 and 21 days, respectively).
FIG. 3. Intestinal sucrase activity. Gerbils received either La1 or
FIG. 1. Shedding of fecal GSA65 antigen. Gerbils received either
placebo daily from 7 days before inoculation with the trophozoites of
La1 (■) or placebo (E) daily from 7 days before inoculation with the
G. intestinalis WB clone C6 (5 ϫ 105 trophozoites per animal). Values
trophozoites of G. intestinalis WB clone C6 (5 ϫ 105 trophozoites per
(arbitrary units [AU] per gram of protein) represent averages from six
animal). Values represent the ratio between GSA65-positive (GSA65
gerbils. Error bars indicate standard deviations. Differences between
ϩ) gerbils and the total number of gerbils at various times after
La1 and placebo groups are statistically significant (P ϭ 0.05 and P ϭ
Giardia infection. The number of animals per group was 18, 12, and 5
0.009 at 14 and 21 days, respectively).