Kamagra enthält Sildenafilcitrat als pharmakologisch aktiven Bestandteil. Dieser hemmt selektiv die Phosphodiesterase-5 und erhöht dadurch die Konzentration von cGMP im Corpus cavernosum. Der Effekt ist zeitlich begrenzt, da die Halbwertszeit von Sildenafil etwa vier Stunden beträgt. In der galenischen Form als Mundgel erfolgt die Resorption besonders rasch, was zu einem schnelleren Wirkeintritt führt. Der Abbau erfolgt überwiegend hepatisch über CYP3A4, wobei ein aktiver Metabolit entsteht, der zur Gesamtwirkung beiträgt. Typische Nebenwirkungen ergeben sich aus der Vasodilatation, darunter leichte Kopfschmerzen und nasale Kongestion. In klinischen Beschreibungen wird kamagra oral jelly im Zusammenhang mit der schnelleren Absorption erwähnt.
Alcohol promotes dopamine release in the human nucleus accumbens
Alcohol Promotes Dopamine Release in the Human Nucleus Accumbens ISABELLE BOILEAU,1 JEAN-MARC ASSAAD,2,4 ROBERT O. PIHL,2 CHAWKI BENKELFAT,3 MARCO LEYTON,3 MIRKO DIKSIC,1 RICHARD E. TREMBLAY,4 AND ALAIN DAGHER1*
1McConnell Brain Imaging Centre, Montreal Neurological Institute, McGill University, Montre´al, QC, Canada
2Department of Psychology, McGill University, Montre´al, QC, Canada
3Department of Psychiatry, McGill University, Montre´al, QC, Canada
4Research Unit on Children’s Psychosocial Maladjustment, Universite´ de Montre´al, Montre´al, QC, Canada
PET; [11C]raclopride; addiction; personality; impulsivity; heart rate;CNS stimulants
Microdialysis experiments in rodents indicate that ethanol promotes
dopamine release predominantly in the nucleus accumbens, a phenomenon that is implicated in the reinforcing effects of drugs of abuse. The aim of the present study was to test the hypothesis in humans that an oral dose of ethanol would lead to dopamine release in the ventral striatum, including the nucleus accumbens. Six healthy subjects underwent two [11C]raclopride PET scans following either alcohol (1 ml/kg) in orange juice or orange juice alone. Subjective mood changes, heart rate, and blood-alcohol levels were monitored throughout the procedure. Personality traits were evaluated using the tridimensional personality questionnaire. PET images were co-registered with MRI and transformed into stereotaxic space. Statistical parametric maps of [11C]raclopride bind- ing potential change were generated. There was a significant reduction in [11C]raclo- pride binding potential bilaterally in the ventral striatum/nucleus accumbens in the alcohol condition compared to the orange juice condition, indicative of increased extra- cellular dopamine. Moreover, the magnitude of the change in [11C]raclopride binding correlated with the alcohol-induced increase in heart rate, which is thought to be a marker of the psychostimulant effects of the drug, and with the personality dimension of impulsiveness. The present study is the first report that, in humans, alcohol promotes dopamine release in the brain, with a preferential effect in the ventral striatum. These findings support the hypothesis that mesolimbic dopamine activation is a common property of abused substances, possibly mediating their reinforcing effects. Synapse 49: 226 –231, 2003. INTRODUCTION
al., 1992; Diana et al., 1992) that are reversed by eth-
Addiction is thought to result in large part from the
anol administration (Weiss et al., 1996). Alcohol is
reinforcing properties of drugs of abuse on brain re-
widely abused by humans; however, no studies have
ward systems, and in particular on mesolimbic dopa-
directly investigated the effect of alcohol consumption
mine (Koob et al., 1998; Wise, 1996). Microdialysis
on the dopamine system. We now present, for the first
studies in rats show that ethanol and other drugs of
time in humans, evidence that alcohol consumed orally
abuse, such as opiates, nicotine, amphetamine, and
promotes dopamine release specifically in the NAc.
cocaine, acutely increase extracellular dopamine levels
We measured dopamine release in response to a sin-
predominantly in the nucleus accumbens (NAc) (Di
gle-dose administration of alcohol using positron emis-
Chiara and Imperato, 1988). The role of NAc dopaminein alcohol self-administration is further supported byreports of changes in consumption following alterations
Contract grant sponsors: the Canadian Institutes for Health Research and the
Fonds de la Recherche en Sante´ du Que´bec.
in mesolimbic dopamine neurotransmission (Rassnick
*Correspondence to: Alain Dagher, Montreal Neurological Institute, 3801 Uni-
et al., 1993; Samson et al., 1993; Nowak et al., 2000),
versity St., Montre´al (QC), Canada, H3A 2B4.
ethanol self-administration into the ventral tegmental
Received 31 January 2003; Accepted 4 April 2003
area (Gatto et al., 1994), and alcohol withdrawal-in-
duced reductions in both dopamine neuron firing andNAc extracellular dopamine concentration (Rossetti et
ALCOHOL PROMOTES DOPAMINE RELEASE IN HUMANS
sion tomography (PET) and the dopamine receptor li-gand [11C]raclopride. We used a two-scan methodbased on evidence in primates, including combinedPET microdialysis studies, that the binding of benz-amides such as [11C]raclopride is sensitive and propor-tional to extracellular dopamine concentration in thestriatum (Endres et al., 1997; Ginovart et al., 1997;Hartvig et al., 1997; Laruelle et al., 1997; Laruelle,2000). This approach has been used in humans to mea-sure the dopamine response to psychostimulants (Car-son et al., 1997; Schlaepfer et al., 1997; Volkow et al.,
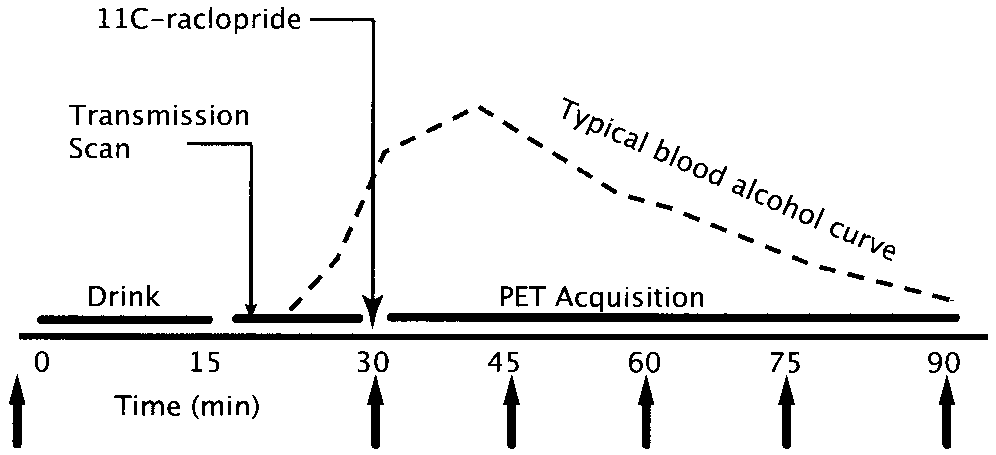
Study design. The vertical arrows indicate the time points
2001) and behavioral tasks (Koepp et al., 1998). Three
of blood sampling, subjective mood assessments, and physiological
recent PET studies have also shown that dopamine
release following amphetamine occurs mostly in theventral striatum and that the amount of dopamine
lected based on previous behavioral experiments show-
released correlates with self-reported behavioral mea-
ing that it was intoxicating but without significant
sures of euphoria or drug wanting (Drevets et al., 2001;
adverse effects in this population. At the end of con-
Leyton et al., 2002; Martinez et al., 2003).
sumption, subjects were immediately positioned in thescanner and a 12-min transmission scan was acquired
MATERIALS AND METHODS
using a 68Ge source for the purpose of attenuation
Seven healthy male nonalcoholic moderate drinkers
correction. Following the transmission scan, and 15
(brief Michigan Alcoholism Screening Test, Pokorny et
min after the end of alcohol consumption, [11C]raclo-
al., 1972), age 22 (Ϯ0.6), were recruited from an exis-
pride 10 mCi was injected as a bolus into the antecu-
tent longitudinal cohort (Tremblay et al., 1994). All
bital vein, after which PET dynamic acquisition (63
subjects who participated had experienced the alcohol
slices, 26 time frames of 60 min total duration) was
dose administered in this study at least twice in a
laboratory setting. Data from one of the seven subjects
Subjects were scanned on the CTI/Siemens ECAT
had to be excluded due to excessive motion during the
HRϩ PET camera with lead septa removed, with in-
scan. All subjects were free of active or past medical or
trinsic resolution 4.8 ϫ 4.8 ϫ 5.6 mm FWHM. Blood
psychiatric illness. Subjects fasted and abstained from
samples, for plasma alcohol measurements, were with-
caffeine or tobacco for a minimum of 4 h before each
drawn from the venous cannula before the initiation of
test session. Five of the six subjects were nonsmokers
drinking, at tracer injection (15 min after finishing
and one was a light smoker (1–2 cigarettes per day).
drinking), and every 15 min thereafter. Subjective ef-
They were also asked to refrain from taking drugs for 7
fects of alcohol, assessed with the Subjective High As-
days and alcohol for 24 h prior to each experimental
sessment Scale (SHAS, Judd et al., 1977; Schuckit et
day. Before each scanning session, subjects underwent
al., 1997), and heart rate were measured prior to alco-
screening for drugs of abuse (Triage Panel for Drugs of
hol consumption and throughout the procedure. The
Abuse, Biosite Diagnostics, San Diego, CA) including
SHAS is a visual analog scale that assesses sensations
alcohol (Alcosensor III intoxicometer, Thomas Instru-
such as feeling high, drunk, and drowsy. In a separate
ments, Montreal, QC). All subjects read and signed a
session, prior to the first scan, all subjects completed
consent form approved by the Research and Ethics
the tridimensional personality questionnaire (TPQ;
Committee of the Montreal Neurological Institute.
Cloninger et al., 1991). This test assesses three dimen-
Subjects participated in two [11C]raclopride PET
sions of personality, including novelty seeking (impul-
scans after consumption of alcohol in orange juice or
sive, excitable, exploratory temperament), which is
orange juice alone (Fig. 1). Subjects were only told
thought to depend in significant part on activity in the
about drink content (alcohol or orange juice alone) at
dopamine pathways (Cloninger, 1994). For the purpose
the beginning of the session, and they did not come into
of anatomical co-registration, subjects also underwent
contact with the drink until the time of consumption.
a 1 ϫ 1 ϫ 1 mm anatomical T -weighted MRI of the
PET data acquisition was performed at the same time
whole brain using a gradient echo pulse sequence
of day (between 14:00 and 16:00) on separate days, 1
(TR ϭ 9.7 ms, TE ϭ 4 ms, flip angle ϭ 12°, FOV ϭ 250,
week apart, and counterbalanced for order of adminis-
tration of alcohol (three out of six received alcohol on
PET frames were summed across time, co-registered
the first day, randomly chosen). Prior to scanning, a
with the corresponding MRI (Woods et al., 1993), and
venous catheter was inserted in the subject’s left arm.
transformed into standardized stereotaxic space (Ta-
Oral consumption of alcohol (1 ml/kg of 95% USP alco-
lairach and Tournoux, 1988) by means of automated
hol over 15 min) or alcohol-free mixture started 30 min
feature-matching to the MNI template (Collins et al.,
prior to tracer injection. The dose of alcohol was se-
1994). Voxelwise [11C]raclopride binding potential (BP)
change in [11C]raclopride BP inducedby an acute oral dose of alcohol (1 ml/kg) in healthy volunteers (n ϭ 6). Colorclusters superimposed on the averageMRI from all subjects depict a signifi-cant change in BP in the ventralstriatum.
was calculated using a simplified reference tissue
session with orange juice that was greater than 2 SD
method (Lammertsma et al., 1996; Gunn et al., 1997) to
from the sample mean, magnitude of heart rate
generate statistical parametric images of the change in
change was analyzed with the nonparametric Wil-
binding (Aston et al., 2000). BP values for each subject
coxon matched pairs test. Maximum change in SHAS
were extracted from regions of interest (ROI) manu-
rating from baseline taken on the same day (⌬max
ally drawn on the co-registered MRI on the left and
[SHAS]) was used to evaluate the subjective effects
right caudate (drawn on transverse slices at Ta-
of alcohol and control drinks. A t-test for paired
lairach-space z coordinate from ϩ2 to ϩ15 mm), pu-
samples was used to determine the difference be-
tamen (ϩ2 to ϩ10 mm), ventral putamen (– 8 to – 4
mm), NAc (– 8 to – 4 mm), and cerebellum, which was
[SHAS] for the control. Stepwise linear regression
used as the reference region. BP values extracted
analysis was used to examine whether percent
from ROI during alcohol and control scans were an-
change in ROI BP could be predicted by changes in
alyzed using a three-way ANOVA for dependent sam-
heart rate, change in SHAS scores, or TPQ person-
ples [Treatment ϫ ROI ϫ hemisphere]. Sphericity
was assessed with the Mauchly test and, when indi-cated, corrections were made with Greenhouse-Gei-
sser adjustments. When appropriate, least signifi-
Screening for drugs of abuse was positive for only one
cant difference t-tests, Bonferroni corrected, were
subject (THC and trace cocaine prior to both scan con-
applied to determine the significance of regional dif-
ditions). Therefore, two different analyses were carried
ferences in BP between the alcohol and orange juice
out, one excluding the data from this subject. In both
conditions. Heart rate during the ascending part of
cases, receptor parametric mapping identified signifi-
the blood alcohol curve was compared to a baseline
cant reductions in [11C]raclopride BP in bilateral ven-
taken just prior to the study session. Since one sub-
tral striatum in the alcohol compared to the alcohol-
ject exhibited a change in heart rate during the test
free condition (Fig. 2). In the statistically generated
ALCOHOL PROMOTES DOPAMINE RELEASE IN HUMANS
DISCUSSION
The observed reduction in [11C]raclopride BP con-
fined to the ventral part of the striatum is indicative ofdopamine release specifically in the NAc and ventralputamen in response to alcohol in humans. The ventralspecificity of the effect is consistent with three other[11C]raclopride PET studies, in which amphetaminewas found to preferentially induce dopamine release inthe ventral striatum in humans (Drevets et al., 2001;Leyton et al., 2002; Martinez et al., 2003). In animals,in vivo microdialysis studies have also shown a propen-sity for alcohol to induce dopamine release in the ven-tral striatum. Di Chiara and Imperato (1988) found
Mean [11C]raclopride BP in the alcohol and control (orange
that ethanol at rewarding doses had an almost 10-fold
juice) conditions. The data are extracted from manually drawn ROI on
greater effect on dopamine release in the NAc than in
each subject’s MRI. Bonferroni corrected pairwise comparisons: †Dif-
the dorsal caudate. Moreover, low doses of ethanol
ference between alcohol and control, P Ͻ 0.001. Error bars representthe SEM.
produce a dose-dependent increase in the firing rate ofA10 dopamine neurons in the ventral tegmental area,
t-map, [11C]raclopride BP values were 16.8 Ϯ 16.3%
which project to the ventral striatum (Gessa et al.,
lower on the test day with alcohol, compared to orange
1985). Activation of A9 dopamine neurons, which
juice (t(5) ϭ 2.54, P ϭ 0.05).
project to the dorsal striatum, only occurs at 5-fold
Analyses of [11C]raclopride BP values in the a priori
defined ROI supported the receptor parametric map-
While a direct pharmacological effect of alcohol could
ping analyses (Fig. 3). A treatment ϫ ROI ϫ hemi-
account for our findings, it is possible that conditioned
sphere ANOVA yielded a main effect of ROI (F(3,15) ϭ
cues and anticipation also played a role in enhancing
42.55, P Ͻ 0.001, Greenhouse-Geisser corrected) and a
dopamine release. In humans, exposure to the odor of
treatment ϫ ROI interaction (F(3,15) ϭ 3.21, P ϭ 0.05).
alcohol leads to autonomic nervous system activity
Bonferroni-corrected pairwise comparisons confirmed
(Stormark et al., 1995), and alcohol-related cues have
that alcohol significantly reduced BP in the NAc (P ϭ
been shown to cause dopamine release in rats previ-
0.003) and ventral putamen (P ϭ 0.001) but not in the
ously trained to self-administer alcohol (Katner and
caudate (P ϭ 0.98) or putamen (P ϭ 0.84). The percent
Weiss, 1999). The subjects in our study only learned
change in [11C]raclopride BP also varied with ROI
whether they would receive alcohol or not when they
(F(3,15) ϭ 13,50, P ϭ 0.001). In both the nucleus ac-
arrived at the lab and they were kept away from the
cumbens (15.0 Ϯ 15.9%) and the ventral putamen
drinks until the start of consumption, thus limiting the
(13.7 Ϯ 16.4%), the percent decreases in [11C]raclopride
potential influence of anticipation in this experiment.
BP were greater than those seen in either the putamen
The dorsal and ventral striatum can be separated
(5.2 Ϯ 17.5%) or caudate nucleus (4.0 Ϯ 16.4%) (P Ͻ
functionally and anatomically (Moore and Bloom, 1978;
Heimer et al., 1982; Haber et al., 2000). Their dopa-
The blood alcohol level reached a mean peak of 18.10
mine innervations originate in different cell groups in
(Ϯ1.4) mmol/L (0.0833 gm %) at 30 min after drinking.
the midbrain and their cortical connections likely ac-
During the expected ascending phase of the blood alco-
count for their different functional roles. The ventral
hol curve (15–30 min post drink) alcohol consumption
striatum, including the NAc and ventral putamen, be-
resulted in small but consistent increases in heart rate
long to the “limbic” cortico-striatal loop that includes
(5.47 Ϯ 6 beats/min; t(5) ϭ 1.85, P ϭ 0.12; 6/6 subjects
the amygdala, hippocampus, orbito-frontal cortex, and
higher during alcohol test, Wilcoxon matched pairs
cingulate cortex, structures involved in emotional be-
test, z ϭ 2.20; P ϭ 0.028) and self-reported feelings of
havior and reward processing. There is much evidence
“high” and “drunkenness” (paired t-test, ⌬
for specific involvement of ventral striatal, or mesolim-
[SHAS] orange juice, P Ͻ 0.01). A
bic, dopamine in the reinforcing effects of addictive
stepwise linear regression showed that impulsiveness,
drugs (Wise, 1996; Koob et al., 1998). It is thought to
one of the subscales on the novelty-seeking dimension
mediate associative learning, whereby drug-related
of the TPQ, and heart rate increase recorded at 30 min
cues acquire incentive value (Di Chiara et al., 1999).
(i.e., during the ascending phase of the blood alcohol
Conditioned place preference, a laboratory test of con-
curve) were the only predictors of BP change in the
ditioned incentive learning, is abolished by lesions or
ventral striatum (r ϭ 0.985; P ϭ 0.005). Neither the
dopamine blockade of the ventral but not dorsal stria-
subjective intoxication measures nor the peak blood
tum (Everitt et al., 1991; Hiroi and White, 1991). Our
alcohol level correlated with the change in [11C]raclo-
finding of dopamine release confined to the ventral
striatum after oral ingestion of an intoxicating dose of
alcohol may therefore account at least in part for its
tiple neurotransmitter systems. In particular, the
SHAS mostly reflects the sedative effects of alcohol
Ethanol most likely acts on dopamine neurons indi-
(Conrod et al., 2001), which are probably not mediated
rectly (Yim et al., 1998). It potentiates GABA-A recep-
tor function (Weiner et al., 1994) to cause inhibition of
In conclusion, we showed that alcohol consumed by
GABAergic interneurons in the substantia nigra re-
mouth in intoxicating doses promotes dopamine re-
ticulata (Mereu and Gessa, 1985), which leads to dis-
lease in the ventral striatum. The observed relation-
inhibition and increased burst firing of dopamine neu-
ship between the magnitude of change in [11C]raclo-
rons (Grace and Bunney, 1985). As stated above, A10
pride BP, personality, and heart rate increase suggests
neurons projecting to the ventral striatum appear to be
that the paradigm we have developed could be used to
more sensitive to these systemic effects of ethanol than
investigate the factors that lead to vulnerability for
A9 dopamine neurons projecting to the dorsal striatum
(Gessa et al., 1985). Opioid peptides may also be in-volved in the dopamine releasing actions of ethanol
REFERENCES
(Acquas et al., 1993; Benjamin et al., 1993; Gonzales etal., 1998).
Acquas E, Meloni M, Di Chiara G. 1993. Blockade of delta-opioid
receptors in the nucleus accumbens prevents ethanol-induced stim-
The level of dopaminergic responsiveness in the NAc
ulation of dopamine release. Eur J Pharmacol 230:239 –241.
has been proposed as a marker of individual vulnera-
Aston JA, Gunn RN, Worsley KJ, Ma Y, Evans AC, Dagher A. 2000.
A statistical method for the analysis of positron emission tomogra-
bility to drug addiction. Numerous studies in rats have
phy neuroreceptor ligand data. Neuroimage 12:245–256.
linked the propensity to self-administer drugs to en-
Benjamin D, Grant ER, Pohorecky LA. 1993. Naltrexone reverses
hanced dopamine release in the NAc in response to
ethanol-induced dopamine release in the nucleus accumbens inawake, freely moving rats. Brain Res 621:137–140.
psychostimulants or stress (e.g., Piazza et al., 1991;
Carson RE, Breier A, de Bartolomeis A, Saunders RC, Su TP, Schmall
Hooks et al., 1992; Zocchi et al., 1998; Marinelli and
B, Der MG, Pickar D, Eckelman WC. 1997. Quantification of am-phetamine-induced changes in [11C]raclopride binding with contin-
White, 2000), or ethanol (Weiss et al., 1993). In light of
uous infusion. J Cereb Blood Flow Metab 17:437– 47.
these studies, our finding of a correlation between the
Cloninger CR. 1994. Temperament and personality. Curr Opin Neu-
change in [11C]raclopride BP in response to alcohol and
Cloninger CR, Sigvardsson S, Bohman M. 1988. Childhood personal-
two variables, the alcohol-induced increase in heart
ity predicts alcohol abuse in young adults. Alcohol Clin Exp Res
rate and the personality trait of impulsiveness, is in-
Cloninger CR, Przybeck TR, Svrakic DM. 1991. The tridimensional
personality questionnaire: U.S. normative data. Psychol Rep 69:
In humans, cardiac response has been hypothesized
Collins DL, Neelin P, Peters TM, Evans AC. 1994. Automatic 3D
to be an index of the psychostimulant properties of
intersubject registration of MR volumetric data in standardized
alcohol and of dopamine activation (Conrod et al.,
Talairach space. J Comput Assist Tomogr 18:192–205.
Conrod PJ, Peterson JB, Pihl RO. 2001. Reliability and validity of
2001), and therefore a marker of vulnerability to ad-
alcohol-induced heart rate increase as a measure of sensitivity to
diction. Our findings, although in a small number of
the stimulant properties of alcohol. Psychopharmacology 157:20 –
subjects, lend support to this theory. Moreover, the
Diana M, Pistis M, Muntoni A, Rossetti ZL, Gessa G . 1992. Marked
personality trait of novelty-seeking, of which impul-
decrease of A10 dopamine neuronal firing during ethanol with-
siveness is one component, has also been linked to both
drawal syndrome in rats. Eur J Pharmacol 221:403– 404.
Di Chiara G, Imperato A. 1988. Drugs abused by humans preferen-
dopamine function (Cloninger, 1994) and to addictive
tially increase synaptic dopamine concentrations in the mesolimbic
propensity. High scores on the novelty-seeking scale of
system of freely moving rats. Proc Natl Acad Sci 85:5274 –5278.
Di Chiara G, Tanda G, Bassareo V, Pontieri F, Acquas E, Fenu S,
the TPQ predict later alcoholism (Cloninger et al.,
Cadoni C, Carboni E. 1999. Drug addiction as a disorder of asso-
1988) as well as relapse-rate in detoxified alcoholics
ciative learning. Role of nucleus accumbens shell/extended amyg-
(Meszaros et al., 1999). Interestingly, in a previous
dala dopamine. Ann NY Acad Sci 877:461– 485.
Drevets WC, Gautier C, Price JC, Kupfer DJ, Kinahan PE, Grace AA,
PET study we found that amphetamine-induced dopa-
Price JL, Mathis CA. 2001. Amphetamine-induced dopamine re-
mine release also targeted the NAc and correlated with
lease in human ventral striatum correlates with euphoria. BiolPsychiatry 49:81–96
novelty-seeking scores (Leyton et al., 2002).
Endres CJ, Kolachana BS, Saunders RC, Su T, Weinberger D, Breier
In the current study, we found no correlation be-
A, Eckelman WC, Carson RE. 1997. Kinetic modeling of [11C]ra-clopride: combined PET-microdialysis studies. J Cereb Blood Flow
tween alcohol-induced NAc dopamine release and sub-
jective measures of intoxication. Previously, we simi-
Everitt BJ, Morris KA, O’Brien A, Robbins TW. 1991. The basolateral
larly found no correlation between amphetamine-
amygdala-ventral striatal system and conditioned place preference:further evidence of limbic-striatal interactions underlying reward-
induced dopamine release and behavioral effects such
related processes. Neuroscience 42:1–18.
as euphoria and excitation (Leyton et al., 2002). The
Gatto GJ, McBride WJ, Murphy JM, Lumeng L, Li TK. 1994. Ethanol
self-infusion into the ventral tegmental area by alcohol-preferring
association between a drug’s euphorigenic quality and
dopamine release has not been established in humans;
Gessa GL, Muntoni F, Collu M, Vargiu L, Mereu G. 1985. Low doses
of ethanol activate dopaminergic neurons in the ventral tegmental
similarly, in rats the behavioral significance of in-
creased DA remains a subject of debate. Our failure to
Ginovart N, Farde L, Halldin C, Swahn CG. 1997. Effect of reserpine-
find a correlation between subjective effects and DA
induced depletion of synaptic dopamine on [11C]raclopride bindingto D2-dopamine receptors in the monkey brain. Synapse 25:321–
release likely reflects the fact that alcohol acts on mul-
ALCOHOL PROMOTES DOPAMINE RELEASE IN HUMANS
Gonzales RA, Weiss F. 1998. Suppression of ethanol-reinforced be-
Moore RY, Bloom FE. 1978. Central catecholamine neuron systems:
haviour by naltrexone is associated with attenuation of the ethanol-
anatomy and physiology of the dopamine systems. Annu Rev Neu-
induced increase in dialysate dopamine levels in the nucleus ac-
cumbens. J Neurosci 18:10663–10671.
Nowak KL, McBride WJ, Lumeng L, Li TK, Murphy JM. 2000. In-
Grace AA, Bunney BS. 1985. Opposing effects of striatonigral feed-
volvement of dopamine D2 autoreceptors in the ventral tegmental
back pathways on midbrain dopamine cell activity. Brain Res 333:
area on alcohol and saccharin intake of the alcohol-preferring P rat.
Alcohol Clin Exp Res 24:476 – 483.
Gunn RN, Lammertsma AA, Hume SP, Cunningham VJ. 1997. Para-
Piazza PV, Rouge-Pont F, Deminiere JM, Kharoubi M, Le Moal M,
metric imaging of ligand-receptor binding in PET using a simplified
Simon H. 1991. Dopaminergic activity is reduced in the prefrontal
reference region model. Neuroimage 6:279 –287.
cortex and increased in the nucleus accumbens of rats predisposed
Haber SN, Fudge JL, McFarland NR. 2000. Striatonigrostriatal path-
to develop amphetamine self-administration. Brain Res 567:169 –
ways in primates form an ascending spiral from the shell to the
dorsolateral striatum. J Neurosci 20:2369 –2382.
Pokorny AD, Miller BA, Kaplan HB. 1972. The brief MAST: a short-
Hartvig P, Torstenson R, Tedroff J, Watanabe Y, Fasth KJ, Bjurling
ened version of the Michigan Alcoholism Screening Test. Am J
P, Langstrom B. 1997. Amphetamine effects on dopamine release
and synthesis rate studied in the rhesus monkey brain by positron
Rassnick S, Stinus L, Koob GF. 1993. The effects of 6-hydroxydopa-
emission tomography. J Neural Transm 104:329 –339.
mine lesions of the nucleus accumbens and the mesolimbic dopa-
Heimer L, Switzer RD, Van Hoesen GW. 1982. Ventral striatum and
mine system on oral self-administration of ethanol in the rat. Brain
ventral pallidum. Components of the motor system? Trends Neuro-
Rossetti ZL, Hmaidan Y, Gessa GL. 1992. Marked inhibition of me-
Hiroi N, White NM. 1991. The amphetamine conditioned place pref-
solimbic dopamine release: a common feature of ethanol, morphine,
erence: differential involvement of dopamine receptor subtypes and
cocaine and amphetamine abstinence in rats. Eur J Pharmacol
two dopaminergic terminal areas. Brain Res 552:141–152.
Hooks MS, Colvin AC, Juncos JL, Justice Jr JB. 1992. Individual
Samson HH, Hodge CW, Tolliver GA, Haraguchi M. 1993. Effect of
differences in basal and cocaine-stimulated extracellular dopamine
dopamine agonists and antagonists on ethanol-reinforced behav-
in the nucleus accumbens using quantitative microdialysis. Brain
iour: the involvement of the nucleus accumbens. Brain Res Bull
Judd LL, Hubbard B, Janowsky DS, Huey LY, Attewell PA. 1977. The
Schlaepfer TE, Pearlson GD, Wong DF, Marenco S, Dannals RF. 1997.
effect of lithium carbonate on affect, mood, and personality of nor-
PET study of competition between intravenous cocaine and [11C]ra-
mal subjects. Arch Gen Psychiatry 34:346 –351.
clopride at dopamine receptors in human subjects. Am J Psychiatry
Katner SN, Weiss F. 1999. Ethanol-associated olfactory stimuli rein-
state ethanol-seeking behavior after extinction and modify extra-
Schuckit MA, Tipp JE, Smith TL, Wiesbeck GA, Kalmijn J. 1997. The
cellular dopamine levels in the nucleus accumbens. Alcohol Clin
relationship between self-rating of the effects of alcohol and alcohol
challenge results in ninety-eight young men. J Stud Alcohol 58:397–
Koepp MJ, Gunn RN, Lawrence AD, Cunningham VJ, Dagher A,
Jones T, Brooks DJ, Bench CJ, Grasby PM. 1998. Evidence for
Stormark KM, Laberg JC, Bjerland T, Nordby H, Hugdahl K. 1995.
striatal dopamine release during a video game. Nature 393:266 –
Autonomic cued reactivity in alcoholics: the effect of olfactory stim-
Koob GF, Sanna PP, Bloom FE. 1998. Neuroscience of addiction.
Talairach J, Tournoux P. 1988. Co-planar stereotaxic atlas of the
Lammertsma AA, Hume SP. 1996. Simplified Reference Tissue Model
Tremblay RE, Pihl RO, Vitaro F, Dobkin PL. 1994. Predicting early
for PET Receptor Studies. Neuroimage 4:153–158.
onset of male antisocial behaviour from preschool behaviour. Arch
Laruelle M. 2000. Imaging synaptic neurotransmission with in vivo
binding competition techniques: a critical review. J Cereb Blood
Volkow ND, Wang G, Fowler JS, Logan J, Gerasimov M, Maynard L,
Ding Y, Gatley SJ, Gifford A, Franceschi D. 2001. Therapeutic doses
Laruelle M, Iyer RN, al-Tikriti MS, Zea-Ponce Y, Malison R, Zoghbi
of oral methylphenidate significantly increase extracellular dopa-
SS, Baldwin RM, Kung HF, Charney DS, Hoffer PB, Innis RB,
mine in the human brain. J Neurosci 21:RC121.
Bradberry CW. 1997. Microdialysis and SPECT measurements of
Weiner JL, Zhang L, Carlen PL. 1994. Potentiation of GABAA-medi-
amphetamine-induced dopamine release in nonhuman primates.
ated synaptic current by ethanol in hippocampal CA1 neurons:
possible role of protein kinase C. J Pharmacol Exp Ther 268:1388 –
Leyton M, Boileau I, Benkelfat C, Diksic M, Baker G, Dagher A. 2002.
Amphetamine-induced increases in extracellular dopamine, drug
Weiss F, Lorang MT, Bloom FE, Koob GF. 1993. Oral alcohol self-
wanting and novelty seeking: A PET/[11C]raclopride study in
administration stimulates dopamine release in the rat nucleus ac-
healthy men. Neuropsychopharmacology 27:1027–1035.
cumbens: genetic and motivational determinants. J Pharmacol Exp
Marinelli M, White FJ. 2000. Enhanced vulnerability to cocaine self-
administration is associated with elevated impulse activity of mid-
Weiss F, Parsons LH, Schulteis G, Hyytia P, Lorang MT, Bloom FE,
brain dopamine neurons. J Neurosci 20:8876 – 8885.
Koob GF . 1996. Ethanol self-administration restores withdrawal-
Martinez D, Slifstein M, Broft A, Mawlawi O, Hwang DR, Huang Y,
associated deficiencies in accumbal dopamine and 5-hydroxytrypta-
Cooper T, Kegeles L, Zarahn E, Abi-Dargham A, Haber SN, Laru-
mine release in dependent rats. J Neurosci 16:3474 –3485.
elle M. 2003. Imaging human mesolimbic dopamine transmission
Wise RA. 1996. Neurobiology of addiction. Curr Opin Neurobiol
with positron emission tomography. II. Amphetamine-induced do-
pamine release in the functional subdivisions of the striatum.
Woods RP, Mazziotta JC, Cherry SR. 1993. MRI-PET registration
J Cereb Blood Flow Metab 23:285–300.
with automated algorithm. J Comput Assist Tomogr 17:536 –546.
Mereu G, Gessa GL. 1985. Low doses of ethanol inhibit the firing of
Yim HJ, Schallert T, Randall PK, Gonzales RA. 1998. Comparison of
neurons in the substantia nigra, pars reticulata: a GABAergic ef-
local and systemic ethanol effects on extracellular dopamine con-
centration in rat nucleus accumbens by microdialysis. Alcohol Clin
Meszaros K, Lenzinger E, Hornik K, Fureder T, Willinger U, Fischer
G, Schonbeck G, Aschauer HN. 1999. The Tridimensional Person-
Zocchi A, Orsini C, Cabib S, Puglisi-Allegra S. 1998. Parallel strain-
ality Questionnaire as a predictor of relapse in detoxified alcohol
dependent effect of amphetamine on locomotor activity and dopa-
dependents. The European Fluvoxamine in Alcoholism Study
mine release in the nucleus accumbens: an in vivo study in mice.
Group. Alcohol Clin Exp Res 23:483– 486.
UM ESTUDO SUPLEMENTAR A VELHA LEI NO NOVO TESTAMENTO OWEN D. OLBRICHT “Agora, com efeito, obteve Jesus ministério tan- digo: até que o céu e a terra passem, nem um i ou to mais excelente, quanto é ele também Mediador um til jamais passará da Lei, até que tudo se de superior aliança instituída com base em supe- riores promessas. Porque, se aquela primeira Essa passage
darunavir 400mg tablets (Prezista®) SMC No. (707/11) Janssen 08 July 2011 The Scottish Medicines Consortium (SMC) has completed its assessment of the above product and advises NHS Boards and Area Drug and Therapeutic Committees (ADTCs) on its use in Scotland. The advice is summarised as follows: ADVICE : following a full submission darunavir (Prezista®) 400mg is accepted f
 ALCOHOL PROMOTES DOPAMINE RELEASE IN HUMANS
sion tomography (PET) and the dopamine receptor li-gand [11C]raclopride. We used a two-scan methodbased on evidence in primates, including combinedPET microdialysis studies, that the binding of benz-amides such as [11C]raclopride is sensitive and propor-tional to extracellular dopamine concentration in thestriatum (Endres et al., 1997; Ginovart et al., 1997;Hartvig et al., 1997; Laruelle et al., 1997; Laruelle,2000). This approach has been used in humans to mea-sure the dopamine response to psychostimulants (Car-son et al., 1997; Schlaepfer et al., 1997; Volkow et al.,
Study design. The vertical arrows indicate the time points
2001) and behavioral tasks (Koepp et al., 1998). Three
of blood sampling, subjective mood assessments, and physiological
recent PET studies have also shown that dopamine
release following amphetamine occurs mostly in theventral striatum and that the amount of dopamine
lected based on previous behavioral experiments show-
released correlates with self-reported behavioral mea-
ing that it was intoxicating but without significant
sures of euphoria or drug wanting (Drevets et al., 2001;
adverse effects in this population. At the end of con-
Leyton et al., 2002; Martinez et al., 2003).
ALCOHOL PROMOTES DOPAMINE RELEASE IN HUMANS
sion tomography (PET) and the dopamine receptor li-gand [11C]raclopride. We used a two-scan methodbased on evidence in primates, including combinedPET microdialysis studies, that the binding of benz-amides such as [11C]raclopride is sensitive and propor-tional to extracellular dopamine concentration in thestriatum (Endres et al., 1997; Ginovart et al., 1997;Hartvig et al., 1997; Laruelle et al., 1997; Laruelle,2000). This approach has been used in humans to mea-sure the dopamine response to psychostimulants (Car-son et al., 1997; Schlaepfer et al., 1997; Volkow et al.,
Study design. The vertical arrows indicate the time points
2001) and behavioral tasks (Koepp et al., 1998). Three
of blood sampling, subjective mood assessments, and physiological
recent PET studies have also shown that dopamine
release following amphetamine occurs mostly in theventral striatum and that the amount of dopamine
lected based on previous behavioral experiments show-
released correlates with self-reported behavioral mea-
ing that it was intoxicating but without significant
sures of euphoria or drug wanting (Drevets et al., 2001;
adverse effects in this population. At the end of con-
Leyton et al., 2002; Martinez et al., 2003).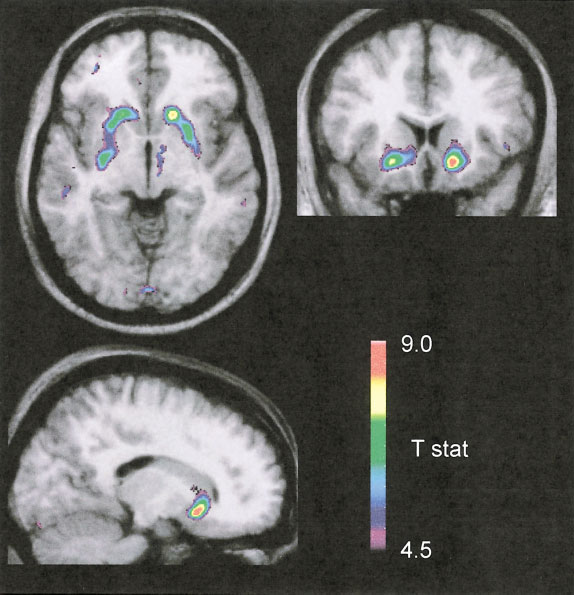 change in [11C]raclopride BP inducedby an acute oral dose of alcohol (1 ml/kg) in healthy volunteers (n ϭ 6). Colorclusters superimposed on the averageMRI from all subjects depict a signifi-cant change in BP in the ventralstriatum.
change in [11C]raclopride BP inducedby an acute oral dose of alcohol (1 ml/kg) in healthy volunteers (n ϭ 6). Colorclusters superimposed on the averageMRI from all subjects depict a signifi-cant change in BP in the ventralstriatum.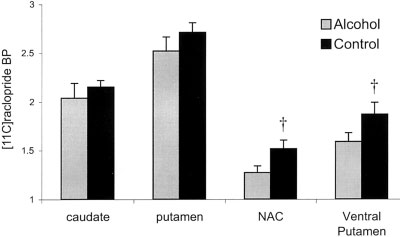 ALCOHOL PROMOTES DOPAMINE RELEASE IN HUMANS
DISCUSSION
ALCOHOL PROMOTES DOPAMINE RELEASE IN HUMANS
DISCUSSION