Kamagra enthält Sildenafilcitrat als pharmakologisch aktiven Bestandteil. Dieser hemmt selektiv die Phosphodiesterase-5 und erhöht dadurch die Konzentration von cGMP im Corpus cavernosum. Der Effekt ist zeitlich begrenzt, da die Halbwertszeit von Sildenafil etwa vier Stunden beträgt. In der galenischen Form als Mundgel erfolgt die Resorption besonders rasch, was zu einem schnelleren Wirkeintritt führt. Der Abbau erfolgt überwiegend hepatisch über CYP3A4, wobei ein aktiver Metabolit entsteht, der zur Gesamtwirkung beiträgt. Typische Nebenwirkungen ergeben sich aus der Vasodilatation, darunter leichte Kopfschmerzen und nasale Kongestion. In klinischen Beschreibungen wird kamagra oral jelly im Zusammenhang mit der schnelleren Absorption erwähnt.
Differential effects of muscle contraction from various body parts on neuromagnetic somatosensory responses
NeuroImage 11, 334–340 (2000) doi:10.1006/nimg.1999.0536, available online at http://www.idealibrary.com on
Differential Effects of Muscle Contraction from Various Body Parts on
Yung-Yang Lin,*,†,‡ Cristina Simo˜es,* Nina Forss,* and Riitta Hari*
*Brain Research Unit, Low Temperature Laboratory, Helsinki University of Technology, P.O. Box 2200, FIN-02015 HUT Espoo, Finland;
†Neurological Institute, Veterans General Hospital-Taipei, Taipei 11217, Taiwan; and ‡Department of Neurology,National Yang-Ming University School of Medicine, Taipei, Taiwan
The early somatosensory-evoked potentials (SEPs) to
We studied eight healthy subjects with a whole-scalp
median nerve stimulation may either increase or de-
306-channel neuromagnetometer to explore the effect
crease during active exploratory finger movements (Cohen
of motor activity from different body parts on somato-
and Starr, 1987; Knecht et al., 1993). Enhancement of SII
sensory responses to left median nerve stimulation.
activation has been observed during finger movements
The stimuli produced clear tactile sensation without
(Huttunen et al., 1996) and sustained contraction of the
any motor movement. In the rest condition, the subject
thenar muscles (Forss and Jousma¨ki, 1998). The interac-
had no task. During contraction conditions, the sub-
tion of different sensory inputs is likely to take place at the
ject had to maintain submaximal isometric contrac-
sensorimotor cortices (Huttunen et al., 1992; Hsieh et al.,tion in masseter, left deltoid, left thenar, or left tibialis
1995). The aim of the present study was to find out how
muscles. Short-latency responses from the primary
isometric contraction of different muscles influences
somatosensory cortex did not change during contrac-
the activation of the somatosensory cortices. tion. Responses from both the right (contralateral) and left second somatosensory cortices (SII) were signifi- cantly enhanced during contraction of the left thenar MATERIALS AND METHODS muscles. Responses from the left SII were significantly enhanced also during contraction of the left deltoid
Somatosensory-evoked magnetic fields (SEFs) were
muscles, but they were decreased during contraction
recorded from eight healthy right-handed subjects (five
of the masseter and left tibialis anterior muscles. This
males, three females; mean age 32 years, range 28–38
study implies that SII activation is modulated by mo-
years). During the recording, the subject was sitting
tor activity and that the effect depends on the topo-
comfortably in a magnetically shielded room with the
graphical proximity of the stimulated and contracted
head leaning against the helmet-shaped neuromagn-
body parts.
2000 Academic Press
etometer. All subjects were experienced in SEF measure-
Key Words: evoked field; magnetoencephalography;
ments, and they were instructed to keep eyes fixed
muscle contraction; somatosensory; human.
forward and to ignore the median nerve stimuli.
The left median nerve was stimulated with 0.3-ms
constant current pulses once every 2 s. The stimulus
INTRODUCTION
intensity was adjusted during the rest condition toproduce a visible twitch of the thumb. We first recorded
Functions of somatosensory and motor cortices are
SEFs with a stimulus intensity clearly exceeding the
closely related. The primary motor cortex receives
motor threshold (6–9 mA, mean 7 mA) to elicit cortical
somatosensory inputs from the thalamus and from the
responses with excellent signal-to-noise ratio. Then,
primary and second somatosensory cortices SI and SII
the intensity of the stimulus was decreased (3–5 mA,
(Jones and Powell, 1968; Zarzecki et al., 1978). On the
mean 4 mA) to produce clear tactile sensation without
other hand, both SI and SII contribute to the pyramidal
any motor movement. SEFs were recorded during five
tract output to modulate motor unit function (Wiesen-
different conditions: (i) rest without voluntary contrac-
danger, 1981). Appropriate integration of the sensorimo-
tion, (ii) contraction of the masseter muscles, (iii)
tor signals is indispensible for the performance of
contraction of the left deltoid muscles, (iv) contraction
of the left thenar muscles, and (v) contraction of the left
Voluntary movement affects the ability to perceive
tibialis anterior muscles. The order of these conditions
stimulation from the moving body part (Coquery et al.,
was randomized over subjects. Before the experiment,
1972; Garland et al., 1972; Angel and Malenka, 1982).
the subject was instructed to bite moderately with
1053-8119/00 $35.00Copyright 2000 by Academic Press
All rights of reproduction in any form reserved.
SEFs of subject 1 to left median nerve stimulation with the stimulus intensity above motor threshold. In each response triplet, the
left traces illustrate signals recorded by the two orthogonal gradiometers (G1 and G2), and the right trace by the magnetometer (M) of a singlesensor unit. The insert shows enlarged responses from the three encircled areas. The vertical line indicates the stimulus onset.
Generators of SEFs and the explanation of the responses by a 3-dipole model in subject 1. The left and middle panels show the
locations of sources superimposed on horizontal and coronal MRI slices, respectively. SI refers to the generator site of N20m, and SIIc and SIIirefer to the contra- and ipsilateral SII, respectively. L, left; R, right. The right panel shows the measured responses (solid lines) from the insertof Fig. 1, superimposed by the wave forms (dotted lines) predicted by the 3-dipole model.
front teeth for masseter activation and to keep a
measured signals, deflections exceeding the prestimu-
comfortable posture during limb muscle contraction.
lus noise level, about 5 fT/(cmͱHz), were first visually
The contractions were monitored throughout the measure-
searched to select the time windows and cortical areas
ments with surface electromyograms and video camera
of interest for further analysis. During these time
recordings. Care was taken to avoid any change in the
windows (from the beginning of the deflection to its
contact of the stimulator with the skin. None of the subjects
return to the baseline level) the magnetic field patterns
complained of muscle fatigue after the experiment.
were first visually surveyed in 2-ms steps to create the
SEFs were recorded with a helmet-shaped 306-
initial guess of the number of active sources within that
channel detector array (Vectorview, Neuromag Ltd,
time period and to estimate the stability of the dipolar
Helsinki, Finland), which comprises 102 identical triple
magnetic field pattern. The equivalent current dipoles
sensor elements. Each sensor element consists of two
(ECDs), best describing the measured data, were found
orthogonal planar gradiometers and one magnetometer
by a least-squares search using subsets of 16–18 chan-
coupled to a multi-SQUID (Superconducting QUantum
nels around the maximum responses. These calcula-
Interference Device) and thus provides three indepen-
tions resulted in the 3-D locations, orientations, and
dent measurements of the magnetic fields. The exact
strengths of the ECDs in a spherical conductor model,
location of the head with respect to the sensors was
which was based on subjects’ MR images. Goodness-
found by measuring the magnetic signals produced by
of-fit of the model was calculated and only ECDs
currents led to four head indicator coils placed at
explaining more than 85% of the field variance at
known sites on the scalp. The locations of the coils with
selected periods of time over a subset of channels were
respect to anatomical landmarks on the head were
determined with a 3-D digitizer to allow alignment of
After identifying the single dipoles, the analysis
the MEG and magnetic resonance (MR) image coordi-nate systems. MR images of the subjects’ brains were
acquired with a 1.5-T Siemens Magnetom scanner.
The signals were bandpass filtered (0.03–200 Hz)
Mean Coordinates (ϮSEM) of the Sources to Left Median
and digitized at 600 Hz. The analysis period of 800 ms
Nerve Stimulation. The Positive x-, y-, and z-Axes Go To-
included a prestimulus baseline of 200 ms, and about
wards the Right Preauricular Point, the Nasion and the
120 responses were averaged for each condition. Ep-
ochs coinciding with signals exceeding 150 µV in the
simultaneously recorded vertical electro-oculogram(EOG) were automatically rejected from the analysis.
The source analysis was based on signals recorded by
the 204 gradiometers. To identify the sources of the
period was extended to the entire measurement epoch
applied in his or her individual multidipole model.
and all channels were taken into account in computing
However, the effect of isometric muscle contraction was
a time-varying multi-dipole model; the strengths of the
quantified only on the SI and SII activations.
previously found ECDs were allowed to change as a
Figure 3 shows the SI and SII source waveforms of
function of time while their locations and orientations
subject 2 during all conditions. The early (20–39 ms) SI
were kept fixed. The validity of the multi-dipole model
responses did not clearly change, but the SII responses
was evaluated by comparing the measured signals with
were differently affected by contraction of various
responses predicted by the model. If signals of any
muscles. In line with a previous study (Forss and
brain region were left inadequately explained by the
Jousma¨ki, 1998), contraction of the left thenar muscles
model, the data were reevaluated for more accurate
enhanced the SII responses bilaterally. Contraction of
estimation of the generator areas. This approach, ex-
the left deltoid muscles also enhanced the SII re-
plained previously in detail (Ha¨ma¨la¨inen et al., 1993),
sponses, although the effect was weaker. In contrast,
has been successfully used in several of our previous
contraction of the masseter and left tibialis anterior
studies (Hari et al., 1993; Forss et al., 1994a; Forss and
muscles slightly decreased the SII responses.
Jousma¨ki, 1998; Nishitani et al., 1999). The individual
Figure 4 shows the mean (ϩSEM; eight subjects)
dipole model derived from responses to stimuli abovemotor threshold was applied to data of all conditions.
Statistical significance of results was tested by Stu-
dent’s paired two-tailed t test.
Figure 1 shows the distribution of SEFs elicited by
stimulation of the left median nerve above motorthreshold in subject 1. Several local maxima of themagnetic field gradients suggest that the stimuli acti-vate several source areas. The earliest deflection N20m,peaking at 20 ms and corresponding to N20 in electricmeasurements, occurred over the right anterior pari-etal cortex (A), followed by P35m at 29 ms. Longer-latency responses peaked over the contralateral tempo-ral region at 122 ms (B) and over the ipsilateraltemporal region at 91 and 140 ms (C).
Figure 2 shows the locations of the source areas
superimposed on the MR images of the same subject. The 3-dipole model (one dipole in SI and two bilaterallyin SII) explained adequately all responses; a singledipole in SI explained satisfactorily both N20m andP35m responses. N20m and P35m were generated inthe postcentral wall of the central fissure. The longer-latency responses were generated bilaterally in theupper lips of the Sylvian fissure in parietal operculum. These generator areas agree with the activation of SIand SII cortices reported in previous magnetoencepha-lographic studies (Hari et al., 1984, 1990; Tiihonen etal., 1989; Forss and Jousma¨ki, 1998). The dipole coordi-nates (Table 1) show on average 5 mm more posteriorSII location in the left than the right hemisphere(P Ͻ 0.05) and are in line with those reported previ-ously (Forss et al., 1994b).
In addition to the activation of the contralateral SI
and of the ipsilateral and contralateral SII, the somato-sensory stimuli also activated the posterior parietal
Strengths of the contralateral SI and the contra- and
cortex in two subjects and the mesial paracentral lobule
ipsilateral SII (SIIc and SIIi) responses of subject 2 as a function of
in 1 subject, as reported previously (Forss et al., 1994a,
time during rest (gray, thin lines) and different contraction conditions
1996). All source areas found in each subject were
Mean (ϩSEM; 8 subjects) change of the dipole strength (difference of amplitude between contraction and rest conditions) of the
N20m, P35m, SIIc, and SIIi responses. Statistical significance *P Յ 0.05; **P Յ 0.01.
change in amplitudes of the SI and SII responses
responses did not differ significantly between the rest
between contraction and rest conditions. N20m
(9.0 Ϯ 2.4 nAm) and P35m (31.1 Ϯ 5.2 nAm) were notsignificantly changed during contraction, whereas re-
DISCUSSION
sponses from the contralateral and ipsilateral SII (SIIcand SIIi, 27.4 Ϯ 3.6 and 30.9 Ϯ 5.1 nAm, respectively)
The present study shows that contraction of various
were 10–60% stronger during contraction of the left
muscles affects differently the activation of the SII
thenar muscles (P Ͻ 0.01 and P Ͻ 0.05, respectively).
cortex. Responses of the left (ipsilateral) SII were
The SIIi responses were significantly enlarged also
enhanced during contraction of the left thenar and
during contraction of the left deltoid muscles, but
deltoid muscles, but decreased during contraction of
reduced during contraction of the masseter and left
the masseter and left tibialis anterior muscles. Similar
tibialis anterior muscles; similar effects were also ob-
changes were observed in the activation of the contralat-
served on the SIIc responses, although those changes
eral SII, although the effect reached statistical signifi-
did not reach statistical significance.
cance only for the thenar muscles. The SI (N20m and
Table 2 shows that the latencies of the SI and SII
P35m) responses did not change significantly withcoincident contraction.
The functional significance of the human SII cortex is
incompletely understood. In monkeys, unilateral SII
lesions impair learning of manual skills (Garcha and
Ettlinger, 1978; Ridley and Ettlinger, 1976). Patientswith selected lesions in SII have been found to have
tactile agnosia (Caselli, 1993), and electrical stimula-
tion of the cortex close to SII area may disturb motor
activity (Penfield and Jasper, 1954). SII has been
suggested to provide an important link between sen-
98.7 Ϯ 7.7 104.2 Ϯ 8.1 98.7 Ϯ 6.8 97.4 Ϯ 5.6 100.3 Ϯ 8.2
sory inputs and motor cortex (Burton, 1986). Our
results indicate that motor activity from various body
the thenar and deltoid muscles also attenuated the SII
parts differentially influences SII activity.
In line with a previous study (Forss and Jousma¨ki,
Thus the modulation of SII activity seems to depend
1998), isometric contraction of the left thenar muscles
on the topographical proximity of the contracting
enhanced SII responses bilaterally. In earlier electric
muscles to the stimulated body part. The effect could
and magnetic studies, the 20-ms responses from the SI
take place either by changes of the synchronicity or of
cortex increased linearly with the increasing stimulus
the number of activated SII neurons. The dependence
intensity up to a certain level (Tsuji et al., 1984;
of SII activation on motor activity at different body
Jousma¨ki and Forss, 1998). Thus the stable amplitude
parts implies spatial tuning (Huttunen et al., 1996),
of N20m between different conditions in the present
which could be helpful for monitoring and correcting
study suggests that the stimulus intensity did not differ
sensorimotor performance. This interpretation is in
significantly between the conditions.
line with monkey studies (Poranen and Hyva¨rinen,
Attention and vigilance are known to modulate more
1982), showing increased responsiveness of SII neu-
strongly responses of the SII than the SI cortex (Hari et
rons during behaviorally important input. al., 1990; Mauguie`re et al., 1997; Mima et al., 1998). Our experienced subjects were familiar with electric
ACKNOWLEDGMENTS
stimuli and tried to ignore them in all conditions. Although we were unable to control their performance
This study was financially supported by Academy of Finland. MR
in this respect, the mesial activation, previously re-
images were obtained from the Department of Radiology of Helsinki
ported to be closely related to voluntary attention to
sensory stimuli (Forss et al., 1996), was observed inonly one subject. Furthermore, the differential effect of
REFERENCES
contraction of different body parts renders the atten-tional factors unlikely to explain our present findings.
Abbruzzese, G., Ratto, S., Favale, E., and Abbruzzese, M. 1981.
Gating effects may occur at cortical or thalamocorti-
Proprioceptive modulation of somatosensory evoked potentialsduring active or passive finger movements. J. Neurol. Neurosurg.
cal sites during active or passive movements, with
Psychiatry 44:942–949.
variable changes in the early (20–45 ms) SEPs (Abbruzz-
Angel, R. W., and Malenka, R. C. 1982. Velocity-dependent suppres-
ese et al., 1981; Cohen and Starr, 1987; Cheron and
sion of cutaneous sensitivity during movement. Exp. Neurol.
Borenstein, 1991). In our study, short-latency (20–39
77:266–274.
ms) responses remained stable during isometric contrac-
Burton, H. 1986. Second somatosensory cortex and related areas. In
tion, suggesting that gating the SI responses requires
Cerebral Cortex (E. G. Jones and A. Peters, Eds.), pp. 31–98. Plenum Press, New York.
changing cutaneous or proprioceptive input.
Caselli, R. J. 1993. Ventrolateral and dorsomedial somatosensory
The SII responses in right-handed subjects are usu-
association cortex damage produces distinct somesthetic syn-
ally stronger in the left than the right hemisphere
dromes in humans. Neurology 43:762–771.
(Forss et al., 1994a), which agrees with anatomical
Cheron, G., and Borenstein, S. 1991. Gating of the early components
studies showing larger operculum in the dominant
of the frontal and parietal somatosensory evoked potentials in
hemisphere (Witelson and Kigar, 1992). Accordingly,
different sensory-motor interference modalities. Electroencepha-
we observed more pronounced interaction between
logr. Clin. Neurophysiol. 80:522–530.
motor activity and SII activation in the left than the
Cohen, L. G., and Starr, A. 1987. Localization, timing and specificity
of gating of somatosensory evoked potentials during active move-
right hemisphere. However, we cannot rule out the
ment in man. Brain 110:451–467.
possibility that this observation was due to the differ-
Coquery, J. M., Coulmance, M., and Leron, M. C. 1972. Modifications
ence between the stimulation sides, because the left
of somaesthetic cortical evoked potentials during active and pas-
hemisphere was always ipsilateral to the stimulation.
sive movements in man. Electroencephalogr. Clin. Neurophysiol.
During finger movements, the SII responses increase
33:269–276.
to stimulation of the median nerve of the same hand,
Forss, N., Hari, R., Salmelin, R., Ahonen, A., Ha¨ma¨la¨inen, M., Kajola,
M., Knuutila, J., and Simola, J. 1994a. Activation of the human
but tend to decrease to stimulation of the other median
posterior parietal cortex by median nerve stimulation. Exp. Brain
nerve (Huttunen et al., 1996). In our study, contraction
Res. 99:309–315.
of the deltoid and thenar muscles, innervated by motor
Forss, N., Salmelin, R., and Hari, R. 1994b. Comparison of somatosen-
cortical areas relatively close to each other, enhanced
sory evoked fields to airpuff and electric stimuli. Electroencepha-
rather similarly the SII responses to wrist stimuli on
logr. Clin. Neurophysiol. 92:510–517.
the same side. In contrast, contraction of the tibialis
Forss, N., Merlet, I., Vanni, S., Ha¨ma¨la¨inen, M., Mauguie`re, F., and
anterior muscles, still on the same side as the stimu-
Hari, R. 1996. Activation of human mesial cortex during somatosen- sory target detection task. Brain Res. 734:229–235.
lated wrist but innervated by motor areas relatively
Forss, N., and Jousma¨ki, V. 1998. Sensorimotor integration in human
remote from the area innervating the thenar muscles,
primary and secondary somatosensory cortices. Brain Res. 781:259–
suppressed the SII responses. Interestingly, contrac-
tion of the masseter muscles, functionally distinct from
Garcha, H. S., and Ettlinger, G. 1978. The effects of unilateral or
bilateral removals of the second somatosensory cortex (area SII): A
Facilitation of somatosensory evoked potentials by exploratory
profound tactile disorder in monkeys. Cortex 14:319–326.
finger movements. Exp. Brain Res. 95:330–338.
Garland, H., Angel, R. W., and Moore, W. E. 1972. Activity of triceps
Mauguie`re, F., Merlet, I., Forss, N., Vanni, S., Jousma¨ki, V., Adeleine,
brachii during voluntary elbow extension: Effect of lidocaine block-
P., and Hari, R. 1997. Activation of a distributed somatosensory
ade of elbow flexors. Exp. Neurol. 37:231–235.
cortical network in the human brain. A dipole modelling study of
Ha¨ma¨la¨inen, M., Hari, R., Ilmoniemi, R. J., Knuutila, J., and
magnetic fields evoked by median nerve stimulation. Part I:
Lounasmaa, O. V. 1993. Magnetoencephalography—Theory, instru-
Location and activation timing of SEF sources. Electroencephalogr.
mentation, and applications to noninvasive studies of the working
Clin. Neurophysiol. 104:281–289.
human brain. Rev. Mod. Phys. 65:413–497.
Mima, T., Nagamine, T., Nakamura, K., and Shibasaki, H. 1998.
Hari, R., Reinikainen, K., Kaukoranta, E., Ha¨ma¨la¨inen, M., Ilmo-
Attention modulates both primary and second somatosensory
niemi, R., Penttinen, A., Salminen, J., and Teszner, D. 1984.
cortical activities in humans: A magnetoencephalographic study. J.
Somatosensory evoked cerebral magnetic fields from SI and SII in
Neurophysiol. 80:2215–2221.
man. Electroencephalogr. Clin. Neurophysiol. 57:254–263.
Nishitani, N., Uutela, K., Shibasaki, H., and Hari, R. 1999. Cortical
Hari, R., Ha¨ma¨la¨inen, H., Ha¨ma¨la¨inen, M., Kekoni, J., Sams, M., and
visuomotor integration during eye pursuit and eye–finger pursuit.
Tiihonen, J. 1990. Separate finger representations at the human
J. Neurosci. 19:2647–2657.
second somatosensory cortex. Neuroscience 37:245–249.
Penfield, W., and Jasper, H. 1954. Epilepsy and the Functional
Hari, R., Karhu, J., Ha¨ma¨la¨inen, M., Knuutila, J., Salonen, O., Sams,
Anatomy of the Human Brain. Little, Brown, Boston.
M., and Vilkman, V. 1993. Functional organization of the human
Poranen, A., and Hyva¨rinen, J. 1982. Effects of attention on multiunit
first and second somatosensory cortices: A neuromagnetic study.
responses to vibration in the somatosensory regions of the mon-
Eur. J. Neurosci. 5:724–734.
key’s brain. Electroencephalogr. Clin. Neurophysiol. 53:525–537.
Hsieh, C.-L., Shima, F., Tobimatsu, S., Sun, S.-J., and Kato, M. 1995.
Ridley, R. M., and Ettlinger, G. 1976. Impaired tactile learning and
The interaction of the somatosensory evoked potentials to simulta-
retention after removals of the second somatic sensory projection
neous finger stimuli in the human central nervous system. A study
cortex (SII) in the monkey. Brain Res. 109:656–660.
using direct recordings. Electroencephalogr. Clin. Neurophysiol.
Tiihonen, J., Hari, R., and Ha¨ma¨la¨inen, M. 1989. Early deflections of
96:135–142.
cerebral magnetic responses to median nerve stimulation. Electro-
Huttunen, J., Ahlfors, S., and Hari, R. 1992. Interaction of afferent
encephalogr. Clin. Neurophysiol. 74:290–296.
impulses in the human primary sensorimotor cortex. Electroen-
Tsuji, S., Shibasaki, H., Kato, M., Kuroiwa, Y., and Shima, F. 1984. cephalogr. Clin. Neurophysiol. 82:176–181.
Subcortical, thalamic and cortical somatosensory evoked poten-
Huttunen, J., Wikstro¨m, H., Korvenoja, A., Seppa¨la¨inen, A.-M.,
tials to median nerve stimulation. Electroencephalogr. Clin. Neuro-
Aronen, H., and Ilmoniemi, R. J. 1996. Significance of the second
physiol. 59:465–476.
somatosensory cortex in sensorimotor integration: Enhancement of
Wiesendanger, M. 1981. Organization of secondary motor areas of
sensory responses during finger movements. Neuroreport 7:1009–
cerebral cortex. In Handbook of Physiology, Section 1, Volume II,Part 2 (J. M. Brookhart and V. B. Mountcastle, Eds.), pp. 1121–
Jones, E. G., and Powell, T. P. S. 1968. The ipsilateral cortical
1147. American Physiological Society, Bethesda.
connexions of the somatic sensory areas in the cat. Brain Res.
Witelson, S. F., and Kigar, D. L. 1992. Sylvian fissure morphology and
9:71–94.
asymmetry in men and women: Bilateral differences in relation to
Jousma¨ki, V., and Forss, N. 1998. Effects of stimulus intensity on
handedness in men. J. Comp. Neurol. 323:326–340.
signals from human somatosensory cortices. Neuroreport 9:3427–
Zarzecki, P., Shinoda, Y., and Asanuma, H. 1978. Projection from area
3a to the motor cortex by neurons activated from group I muscle
Knecht, S., Kunesch, E., Buchner, H., and Freund, H. J. 1993.
afferents. Exp. Brain Res. 33:269–282.
List of publications, Laurent M. Haegeli 1. Original papers (OP): Arentz T, Weber R, Jander N, Bürkle G, von Rosenthal J, Blum T, Stockinger J, Haegeli LM , Neumann FJ, Kalusche D. Pulmonary haemodynamics at rest and during exercise in patients with significant pulmonary vein stenosis after radiofrequency catheter ablation for drug resistant atrial fibrillation. Eur Heart J 2005 Jul
Palliative Care Guidelines: Last days of life Palliative Care in the last days of life Introduction This guideline is an aid to clinical decision-making and good practice in person-centred care for patients who are deteriorating and at risk of dying. The patient may have a new life-limiting condition, and / or have one or more advanced illnesses. A decision will have been made that transf

 NeuroImage 11, 334–340 (2000)
NeuroImage 11, 334–340 (2000)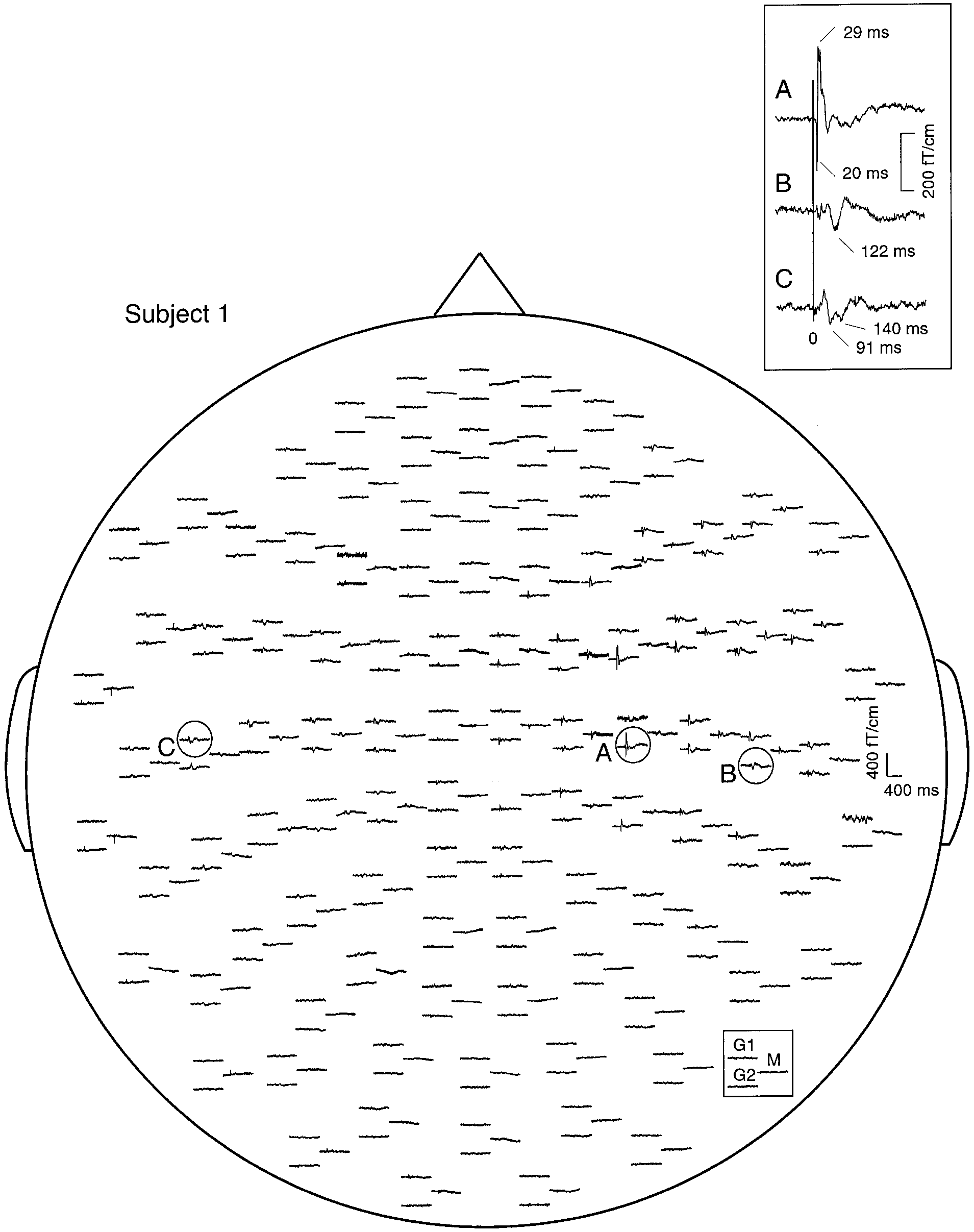 SEFs of subject 1 to left median nerve stimulation with the stimulus intensity above motor threshold. In each response triplet, the
left traces illustrate signals recorded by the two orthogonal gradiometers (G1 and G2), and the right trace by the magnetometer (M) of a singlesensor unit. The insert shows enlarged responses from the three encircled areas. The vertical line indicates the stimulus onset.
SEFs of subject 1 to left median nerve stimulation with the stimulus intensity above motor threshold. In each response triplet, the
left traces illustrate signals recorded by the two orthogonal gradiometers (G1 and G2), and the right trace by the magnetometer (M) of a singlesensor unit. The insert shows enlarged responses from the three encircled areas. The vertical line indicates the stimulus onset.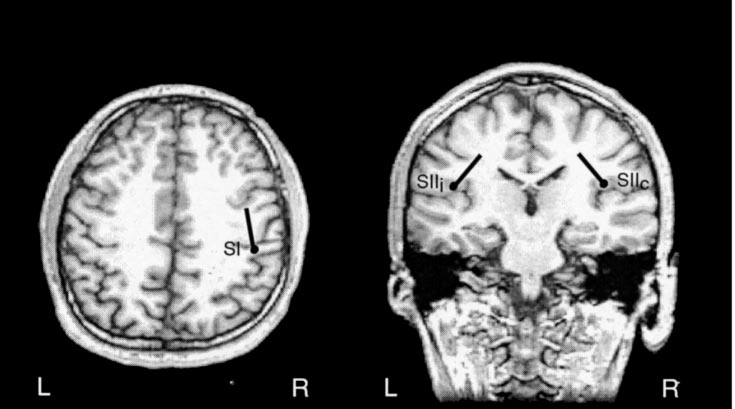
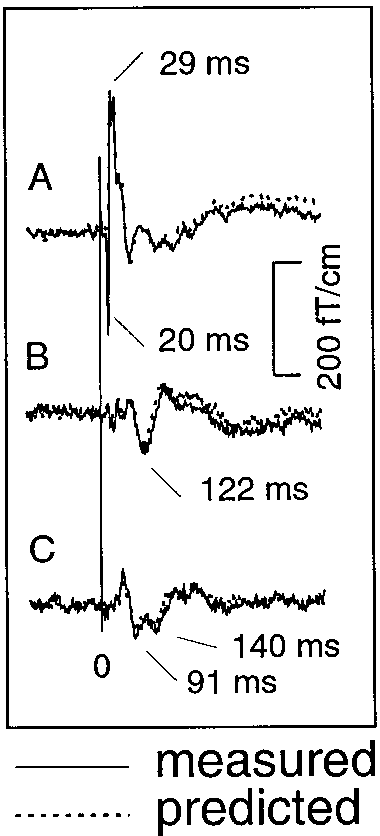 Generators of SEFs and the explanation of the responses by a 3-dipole model in subject 1. The left and middle panels show the
locations of sources superimposed on horizontal and coronal MRI slices, respectively. SI refers to the generator site of N20m, and SIIc and SIIirefer to the contra- and ipsilateral SII, respectively. L, left; R, right. The right panel shows the measured responses (solid lines) from the insertof Fig. 1, superimposed by the wave forms (dotted lines) predicted by the 3-dipole model.
Generators of SEFs and the explanation of the responses by a 3-dipole model in subject 1. The left and middle panels show the
locations of sources superimposed on horizontal and coronal MRI slices, respectively. SI refers to the generator site of N20m, and SIIc and SIIirefer to the contra- and ipsilateral SII, respectively. L, left; R, right. The right panel shows the measured responses (solid lines) from the insertof Fig. 1, superimposed by the wave forms (dotted lines) predicted by the 3-dipole model.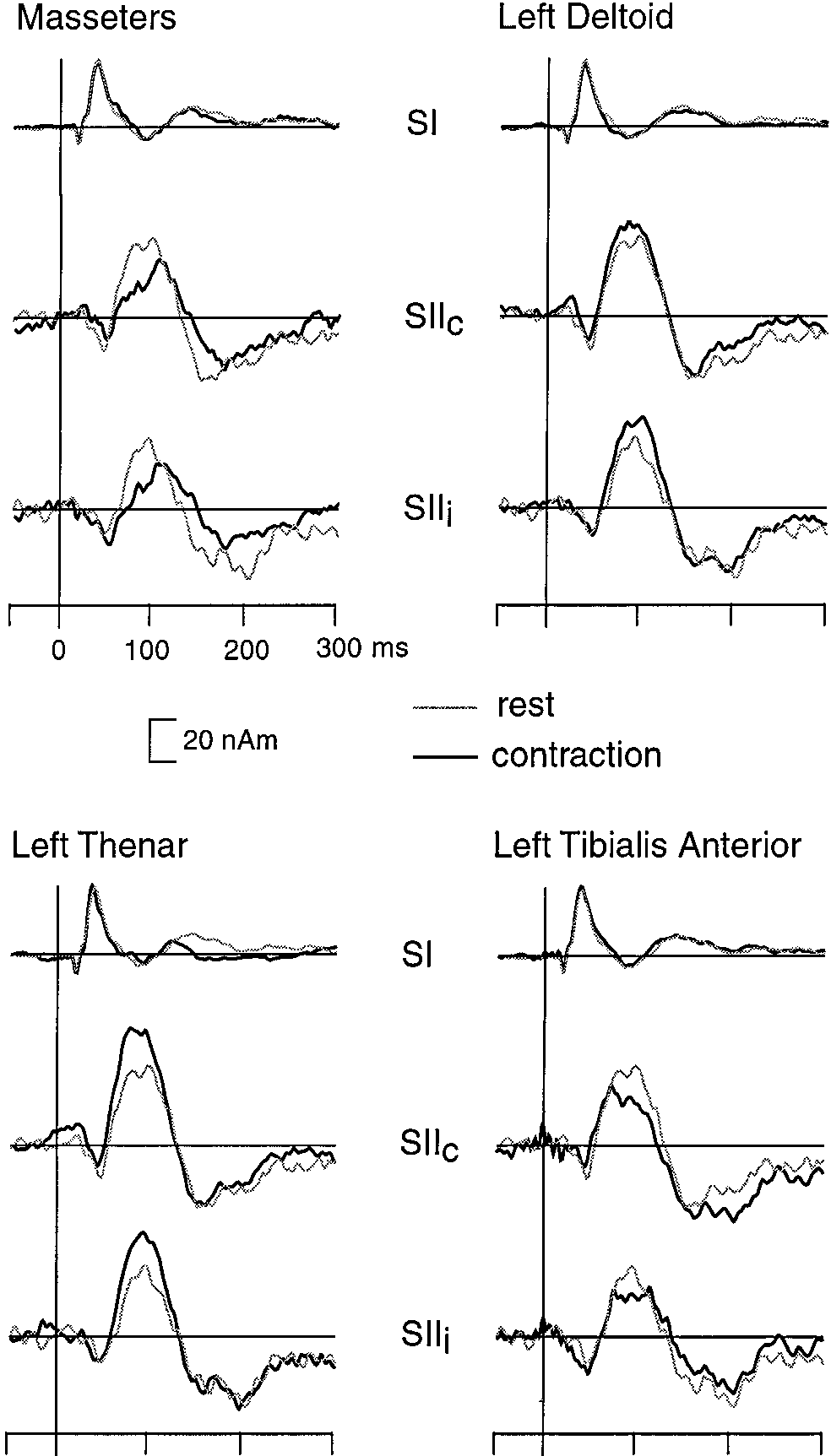 period was extended to the entire measurement epoch
applied in his or her individual multidipole model.
period was extended to the entire measurement epoch
applied in his or her individual multidipole model.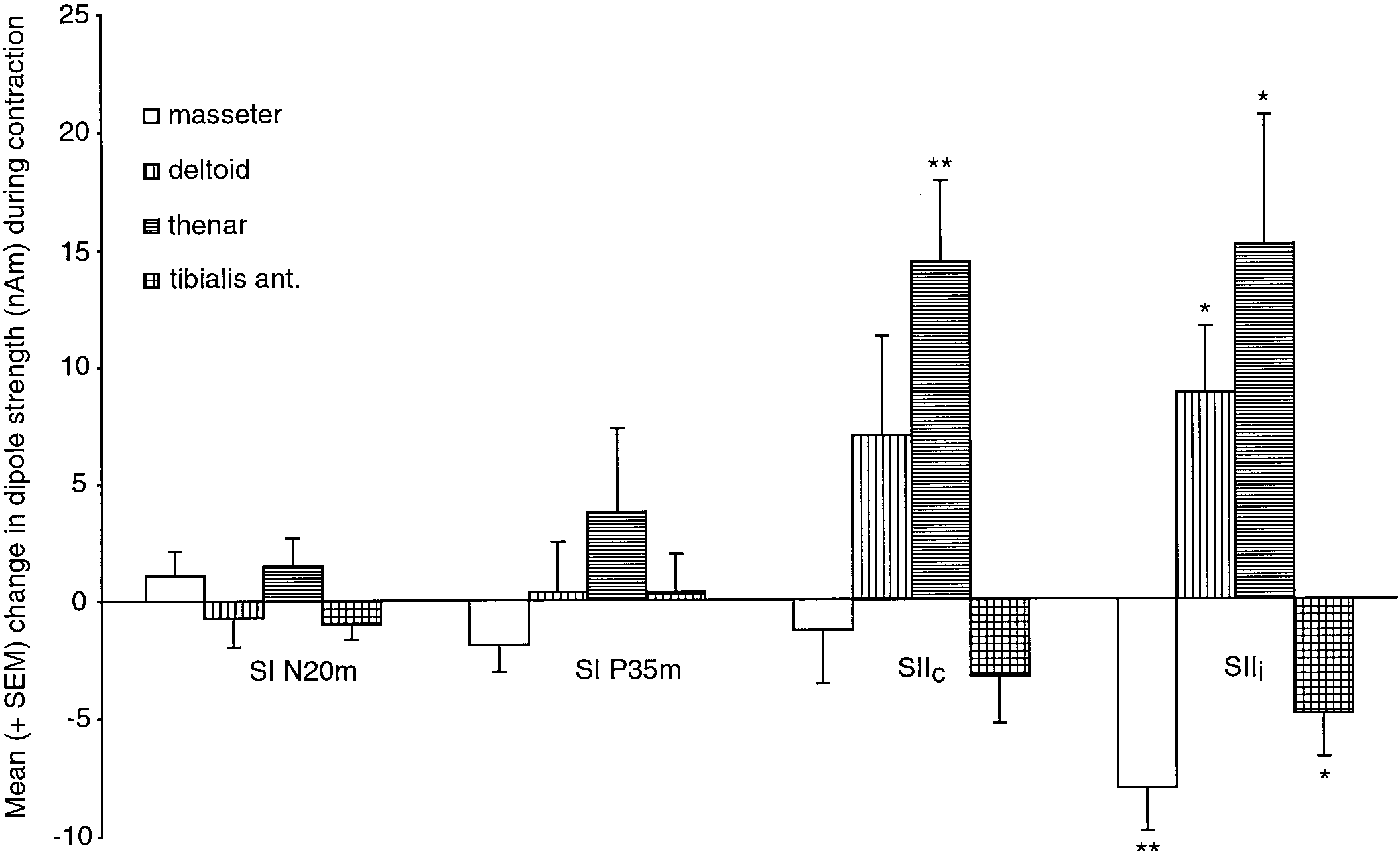 Mean (ϩSEM; 8 subjects) change of the dipole strength (difference of amplitude between contraction and rest conditions) of the
N20m, P35m, SIIc, and SIIi responses. Statistical significance *P Յ 0.05; **P Յ 0.01.
Mean (ϩSEM; 8 subjects) change of the dipole strength (difference of amplitude between contraction and rest conditions) of the
N20m, P35m, SIIc, and SIIi responses. Statistical significance *P Յ 0.05; **P Յ 0.01.