Kamagra enthält Sildenafilcitrat als pharmakologisch aktiven Bestandteil. Dieser hemmt selektiv die Phosphodiesterase-5 und erhöht dadurch die Konzentration von cGMP im Corpus cavernosum. Der Effekt ist zeitlich begrenzt, da die Halbwertszeit von Sildenafil etwa vier Stunden beträgt. In der galenischen Form als Mundgel erfolgt die Resorption besonders rasch, was zu einem schnelleren Wirkeintritt führt. Der Abbau erfolgt überwiegend hepatisch über CYP3A4, wobei ein aktiver Metabolit entsteht, der zur Gesamtwirkung beiträgt. Typische Nebenwirkungen ergeben sich aus der Vasodilatation, darunter leichte Kopfschmerzen und nasale Kongestion. In klinischen Beschreibungen wird kamagra oral jelly im Zusammenhang mit der schnelleren Absorption erwähnt.
Reboundhealth.net
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Nov. 2006, p. 3519–3528
0066-4804/06/$08.00ϩ0 doi:10.1128/AAC.00545-06Copyright 2006, American Society for Microbiology. All Rights Reserved. MINIREVIEW
Impact of Melanin on Microbial Virulence and Clinical Resistance
Joshua D. Nosanchuk1* and Arturo Casadevall1,2
Department of Medicine, Division of Infectious Diseases,1 and Microbiology and Immunology,2
Albert Einstein College of Medicine, Bronx, New York
Melanins are negatively charged, hydrophobic pigments of
cumstances is uncertain, such as melanin in the neurons of the
high molecular weight (54, 88, 95, 139) that are composed of
substantia nigra in the human brain (147, 148).
polymerized phenolic and/or indolic compounds (Fig. 1) (45,
In mammals, melanin synthesis is catalyzed by a tyrosinase
128). Melanins are produced by organisms in all biological
(114). In contrast, microbes generally synthesize melanin via
kingdoms, including a wide variety of pathogenic bacteria,
various phenoloxidases (such as tyrosinases, laccases, or cata-
fungi, and helminths (reviewed in reference 90). Remarkably
cholases) and/or the polyketide synthase pathway (reviewed in
little is known about the structures of melanins, despite their
reference 138). Melanins generated from 3,4-dihydroxyphenyal-
abundance in the global biomass. This is due to the inability of
anine (DOPA) by phenoloxidases are referred to as eumela-
current biochemical and biophysical techniques to provide a
nins, which are generally black or brown. Yellow or reddish
definitive chemical structure, because these complex polymers
melanins are called pheomelanins and incorporate cysteine
are amorphous, insoluble, and not amenable to either solution
with DOPA. Brownish melanins derived from homogentisic
or crystallographic structural studies. Consequently, our infor-
acid by tyrosinases are called pyomelanins (144). Melanins
mation on the structure of melanin is derived from the analysis
formed from acetate via the polyketide synthase pathway are
of their degradation products and spectroscopic analysis of the
typically black or brown and are referred to as dihydroxynaph-
melanin polymer (128). Characteristically, melanins are dark in
color, insoluble in aqueous or organic fluids, resistant to con-
Melanin synthesis has been associated with virulence for a
centrated acid, and susceptible to bleaching by oxidizing agents
variety of pathogenic microbes. Melanin is believed to contrib-
(17, 87, 103). Methods for partial chemical degradation of
ute to microbial virulence by reducing a pathogen’s suscepti-
melanin followed by high-pressure liquid chromatographic mi-
bility to killing by host antimicrobial mechanisms and by influ-
croanalysis have been developed and are useful for the char-
encing the host immune response to infection. Consequently,
acterization of specific types of melanin (128, 129). An oper-
melanin and melanin synthesis pathways are potential targets
ational definition for a pigment as a melanin can be provided
for antimicrobial drug discovery. Interestingly, the drug-bind-
by electron spin resonance characteristics, since these pig-
ing properties of both host and microbial melanins could in-
ments uniquely are stable organic free radicals (29).
fluence the outcome of antimicrobial therapy.
Many diverse functions have been attributed to melanins.
This review discusses the impact of melanin production on
Melanins can serve as energy transducers and affect cellular
microbial survival in the environment and during infection, on
integrity (reviewed in reference 48). Melanin is also used for
host immune responses, and on the efficacies of antimicrobial
sexual display and camouflage. For instance, the coloration in
compounds. The capacity for melanin to bind to diverse com-
black and red hair arises from melanin (18). An example in
pounds can affect the testing of antimicrobial drugs and reduce
which melanin is used for camouflage is the release of ink, a
the activity of antimicrobial therapy.
suspension of melanin particles, by the cuttlefish (Sepia offici-nalis) in response to danger (34). Melanin plays a major role inthe innate immune system of insects, which synthesize the
MELANINS CONFER A SURVIVAL ADVANTAGE TO
polymer to damage and entomb microbial intruders (85, 104). ENVIRONMENTAL MICROBES
In insects, invading microbes activate a prophenoloxidase inthe hemolymph, resulting in the encasement of the bacterial,
Melanin synthesis in free-living microbes is likely to provide
protozoal, or fungal pathogen in melanin (78). Melanins in
a survival advantage in the environment (Table 1) (117). This
melanocytes in skin provide protection against sunlight and are
hypothesis is based on the fact that many fungi constitutively
also believed to contribute to the resistance of melanoma to
synthesize melanin, and even facultative melanotic microbes
therapeutic radiation (47). The role of melanin in other cir-
like Cryptococcus neoformans are melanized in soils (94). Mel-anin production in C. neoformans is associated with increasedsurvival after ingestion by environmental amoeboid (118) ornematode (84) predators. Environmental predators often pro-duce hydrolytic enzymes to digest microbes, and melanized C.
* Corresponding author. Mailing address: Albert Einstein College of
neoformans cells are significantly less susceptible to cell wall-
Medicine, 1300 Morris Park Avenue, Bronx, NY 10461. Phone: (718)430-3766. Fax: (718) 430-8968. E-mail: nosanchu@aecom.yu.edu.
degrading enzymes than nonmelanized cells (109). Melanin
FIG. 1. Chemical structures of pheomelanin (A) and eumelanin (B) oligomers.
production in diverse environmental melanotic molds has been
for the growth of black fungi in the highly contaminated Cher-
associated with reduced cellular susceptibility to enzymatic
nobyl Reactor No. 4 (83). The pigment significantly contributes
degradation (reviewed in reference 90). The mechanism of
to the ability of C. neoformans to withstand extremes in heat
action for resistance to enzymatic hydrolysis is unclear but may
and cold (108). In fungal plant pathogens, melanization of
involve sequestration of the enzymes on melanin or may occur
appressorium allows a cell to maintain integrity while gener-
by steric hindrance (54). Additional evidence supporting a
ating pressures in excess of 80 bar to facilitate the penetration
protective role for melanin is provided by the fact that addition
peg’s entry into a plant cell (reviewed in reference 90).
of synthetic melanin to suspensions of Aspergillus nidulans re-
Melanins are able to bind to the heavy metals that are
sults in significant inhibition of the hydrolytic activity of glu-
routinely found in the environment (35, 105, 153). The car-
canase-chitinase on the fungus (67).
boxyl, phenolic, hydroxyl, and amine groups on melanin pro-
Melanins confer resistance to UV light by absorbing a broad
vide numerous potential binding/biosorption sites for metal
range of the electromagnetic spectrum and preventing photo-
ions (reviewed in reference 35). Melanized C. neoformans cells
induced damage (48). Consequently, melanins are used com-
are more resistant to killing by silver nitrate, a compound
mercially in photoprotective creams and eye glasses. Melanin
highly toxic to bacteria and fungi, than nonmelanized cells
protects several fungal and bacterial species from UV, solar, or
(42). That study demonstrated that melanin chelated the silver
gamma radiation (reviewed in reference 90). Increased mela-
compound (42). Although other fungal melanins bind to met-
nin production is associated with the greater resistance of
als (reviewed in reference 35), a protective role for metal
pigmented fungi to radiation (127, 149, 150). The protective
binding has not been demonstrated in other microbes. How-
properties of melanin against radiation injury could account
ever, the evidence from C. neoformans suggests that the utilityof metals as antimicrobial drugs against melanin-producingorganisms may be lower than that against non-melanin-pro-
TABLE 1. Melanization protects pathogenic fungi from
MELANIN PROTECTS MICROBES FROM HOST DEFENSES
The ability of melanin to protect microbes from host de-
fenses is relevant to antimicrobial therapy because the clinical
efficacies of some antimicrobial drugs are complemented by
host immune defenses. Melanin has been shown to interfere
with numerous host defense mechanisms (Table 2). Melanized
C. neoformans yeast cells manifest increased resistance to
phagocytosis in vitro and in vivo (82, 130). In macrophage-like
cell lines, the phagocytic index for melanized Paracoccidioidesbrasiliensis yeast cells was half that for the nonmelanized cells
(21). Since melanins are charged polymers (139), their pres-
TABLE 2. Melanin production protects pathogenic fungi against
dermatitidis do not reduce phagocytosis or protect against
oxidative burst or killing by neutrophils (115).
Melanins are highly effective scavengers of free radicals
(116) and have electron transfer properties (41). Electrontransfer from free radical species generated in solution to
melanin derived from C. neoformans has been demonstrated
by electron spin resonance spectroscopy (131); and similar
spectra have been generated with melanins from Histoplasmacapsulatum, S. schenckii, P. brasiliensis, and Pneumocystis spp. (reviewed in reference 90). Also, C. neoformans melanin is
involved in the reduction of Fe3ϩ to Fe2ϩ (97) and can facil-
itate redox cycling through the exportation of electrons to form
extracellular Fe2ϩ, which maintains the reducing capacity of
extracellular redox buffers (56). Melanized C. neoformans cells
are also less susceptible to the toxic effects of microbicidal
peptides than nonmelanized cells (26). The mechanism of ac-
tion in this case appears to be adsorption of the microbicidal
peptide such that it interferes with the peptide reaching its
EVIDENCE THAT MELANINS BIND a Magnitude indicates the maximal percent increase in protection afforded to
TO DRUGS IN VITRO
the organism by the production of melanin compared to that afforded to cellsdeficient in melanin. Isotherm analysis of adsorption of drugs by melanin. Mel-
Represents attachment rather than ingestion of fungal cells. c As measured by mitochondrial damage rather than numbers of CFU.
anins bind to chemically diverse compounds (62, 70). Thebinding of gentamicin, methotrexate, and chlorpromazine tomelanins has recently been revisited by using isotherm binding
equations to characterize the adsorption of the drugs to syn-
ence in the cell wall of C. neoformans can alter the fungal cell
thetic and Sepia officinalis melanins (14). Although there were
surface charge (88), and this may contribute to inhibition of
significant variations in adsorption, each drug bound to mela-
phagocytosis. Melanization increased the cellular negative
nin. More gentamicin than the other drugs was bound by syn-
charge by 3 to 33% in nine different encapsulated strains and
thetic melanin. By the best-fit Freundlich equation for genta-
by 86% in an acapsular strain (88). In addition to reducing
micin [q ϭ q (KC)1/n dm3 · gϪ1, where q is the amount
ingestion, melanization protects C. neoformans against killing
absorbed [mmol · gϪ1], q is the adsorption capacity, K is the
by macrophages (130). Similarly, melanin production in Fon-
energy of absorption, C is the equilibrium solution concentra-
secaea pedrosoi (20), Sporothrix schenckii (107), and Exophiala
tion of solute, and the heterogeneity index 1/n is between 0 and
spp. (33, 99, 115) enhances resistance to killing by phagocytic
1], the quantity of gentamicin absorbed with synthetic melanin
cells. Melanin in pigmented C. neoformans yeast cells can pro-
was 0.49 dm3 · gϪ1, whereas 0.061 dm3 · gϪ1 of methotrexate
duce complex immunomodulatory effects that contribute to
was bound. Equilibrium isotherms for the interaction of gen-
virulence by eliciting changes in the host cytokine/chemokine
tamicin with melanin revealed diverse interactions between the
response (50, 82). In particular, melanized yeast resulted in
drug and melanin. In contrast, methotrexate appeared to bind
higher levels of interleukin-4 and monocyte chemoattractant
to specific homogeneous sites. The data from that study (14)
protein 1 and increased the numbers of pulmonary leukocytes
demonstrated the significant capacity of melanin to interact
early after infection (82). F. pedrosoi melanin also activates
with drugs, since its adsorption capacity was comparable to
those of other absorbers, like medicinal activated charcoal.
Melanization protects fungi, such as C. neoformans, Aspergil-Scatchard plot analysis of drug binding by melanin. A Scat- lus spp., and S. schenckii, and bacteria, such as Proteus mirabilis
chard plot-type analysis of drug binding to melanin by the use
and Burkholderia cepacia, against injury secondary to nitrogen-
of radiolabeled compounds has also demonstrated the pres-
or oxygen-derived radical attack (reviewed in reference 90). F.
ence of heterologous binding sites. There are at least two
pedrosoi melanin significantly inhibits nitric oxide production
classes of binding sites on synthetic DOPA melanin for the
by macrophages, which affects the pathogenesis of chronic
aminoglycoside antibiotics gentamicin (141) and kanamycin
chromoblastomycosis (10). The melanized F. pedrosoi cells re-
(142). For kanamycin, the association constants for the strong
duced the production of nitric oxide by approximately 50%
and weak binding sites were 3 ϫ 105 MϪ1 and 4 ϫ 103 MϪ1,
compared to the amount produced by nonstimulated murine
respectively, and 0.64 M kanamycin was required to saturate
peritoneal macrophages and, similarly, suppressed nitric oxide
the binding sites in 1 mg melanin (142). Scatchard plot-type
production in macrophages stimulated by interferon gamma
analyses with melanins have also revealed high- and low-affin-
and lipopolysaccharide. Interestingly, melanin is not the only
ity binding sites for cocaine (61, 102); amphetamines (6, 43);
pigment that can modify the effects of oxidative attacks by host
donarubicin (120); and the antiarrhythmics quinidine, disopyr-
cells, since carotenoids in Staphylococcus aureus similarly func-
amide, and metoprolol (16). More recently, high-resolution
tion as antioxidants (75). However, carotenoids in Exophiala
magic angle spinning nuclear magnetic resonance spectroscopy
revealed highly specific melanin-binding sites for iodobenz-amides (11), which can be exploited to diagnose and stagemelanoma by using radiolabeled drug. Absorption studies with antifungals. Two methods have
been used to establish that melanin binds to amphotericin Band caspofungin. First, the ability of melanin produced by C. neoformans and synthetic melanin to bind to these antifungaldrugs was inferred from experiments whereby melanins wereincubated with various compounds and then the antifungalactivity of the solution was determined (52, 124, 125). Forthose studies, melanin particles were removed by centrifuga-tion prior to the testing of the antifungal drug solutions in MICand time-kill studies. Incubation of amphotericin B and caspo-fungin with melanin significantly reduced their antifungalactivities for C. neoformans. In contrast, incubation of itra-conazole, fluconazole, or flucytosine with melanin had no ef-fect on their antifungal activities. One study showed a 16-foldincrease in the MIC of amphotericin B after adsorption of thedrug with 1 ϫ 107 melanin particles derived from pigmented C. neoformans prior to MIC testing (52). Furthermore, the in-crease in MIC correlated with the amount of melanin particles
added to the antifungal drug solution. Time-kill assays alsodemonstrated that the addition of melanin particles to ampho-
FIG. 2. The pathogenic yeast Cryptococcus neoformans. (A) India
tericin B or caspofungin significantly reduced their toxicities
ink preparation showing a budding C. neoformans yeast cell with alarge polysaccharide capsule surrounding the cell bodies. Bar, 5 m.
for C. neoformans (124). For example, 71% of yeast cells sur-
(B) A melanin “ghost,” a melanin particle isolated from C. neoformans
vived exposure to 2ϫ the MIC of amphotericin B preincubated
grown for 10 days in the presence of L-dopa by serial treatment of the
with synthetic melanin, whereas the rate of survival was 8% for
yeast with enzymes, denaturant, chloroform, and hot acid. Bar, 2 m.
the cells exposed to amphotericin B not incubated with mela-
(C) Transmission electron micrograph of a cross-section of a C. neo-formans “ghost” showing that the particle is formed of concentric
nin. Similarly, 79% of yeast cells exposed to caspofungin pre-
layers of melanin. Bar, 1 m. (D) Depiction of the melanin granules
incubated with melanin survived exposure to drug, whereas the
comprising the melanin layers, demonstrating how the packing of the
rate of survival was 11% for cells incubated with native caspo-
granules results in pores that obstruct the passage of large molecules,
fungin. In contrast, incubation of azoles or flucytosine with
such as amphotericin B or caspofungin. Obstruction of antifungal
melanin did not affect their MICs for C. neoformans or the
molecules can occur by virtue of a reduced melanin pore size ormelanin binding. Panel D is based on data from reference 27.
abilities of the drugs to kill C. neoformans.
Antifungal drug binding to fungal melanin was also inferred
by the finding that the elemental composition of melanin waschanged after incubation with antifungal drugs. Incubation of
of melanin particles arranged in a concentric manner. The
amphotericin B or caspofungin with melanin altered the mel-
thickness of the layer appears to depend on cell age, such that
anin C:N:O ratio, consistent with drug absorption (124). In
older cells may be significantly less susceptible to melanin-
contrast, no change in the melanin elemental composition was
binding antifungal drugs than younger cells. In order for the
observed following the incubation of melanin with flucytosine,
fungal cells to survive, the polymer layers composed of gran-
voriconazole, fluconazole, or itraconazole (124, 125). Since
ules cannot completely exclude nutrients. Nuclear magnetic
melanin is located in the cell wall, these data suggest and are
resonance cryoporometry revealed that melanin ghosts contain
consistent with a mechanism of acquired resistance, whereby
pores with diameters between 1 and 4 nm, in addition to a
fungal melanin binds to amphotericin B and caspofungin and
small number of pores with diameters nearly 30 nm. The pore
prevents them from reaching their target sites.
size decreases with the age of the yeast cell. Importantly, cryo-
Effect of melanin on the porosity of the microbial cell wall.
porometry studies with melanin-binding antibody show that
Analysis of the microstructure of cell wall-associated melanin
the larger pores appear to be internal to the smaller pores. In
in C. neoformans has provided new insights into the potential
another study, the porosity of melanized cryptococcal cell walls
of this polymer to interfere with antifungal drug activity (27).
was evaluated by elution of graded dextrans, and similar results
Cell wall-associated melanin is composed of discrete granules
were described (57). These findings suggest that the tight
of roughly uniform dimensions (Fig. 2). This is significant,
spaces between melanin granules may prevent or slow the
because a granular arrangement would significantly increase
entry of large drugs, such as amphotericin B (molecular mass,
the surface area available for binding to certain types of drugs.
924 g/mol) and caspofungin (molecular mass, 1,093.5 g/mol),
Atomic force microscopy and transmission electron micros-
into pigmented cells. This may be particularly significant for
copy revealed that the melanin particles range in size from 40
amphotericin B, since this drug tends to form large aggregates
to Ͼ100 nm, with an average particle diameter of 76 nm, which
in solution (68). In contrast, azoles and flucytosine have sig-
is similar to the results for mammalian melanin and melanin
nificantly smaller molecular masses, and for these compounds,
from S. officinalis (27). Transmission electron microscopy re-
melanization does not reduce fungal cell susceptibility. Hence,
vealed that cell wall melanin is composed of two to five layers
melanin in the cell wall may also reduce the susceptibilities to
certain drugs by inhibiting their diffusion into the cell body and
The interaction of these chemotherapeutics with melanin may
provide greater opportunity for the binding of drug by melanin.
be responsible for decreased wound healing and for an unsat-isfactory response of the tumor to the medication. However,many melanin-binding drugs commonly used in clinical prac-
BINDING OF COMPOUNDS BY MELANIN
tice, such as beta-blockers, benzodiazepines, and rifamycins,
IN HUMANS IN VIVO
bind to melanin in vitro, without apparent adverse effects in
The binding of drugs to host melanin can damage certain
tissues, and drug-melanin interactions have been implicated in
Aminoglycosides are positively charged at physiological pH
the pathogenesis of several diseases, such as Parkinson’s dis-
and have a relatively high molecular weight, which limits their
ease. In Parkinson’s disease there is a loss of pigment in the
penetration into tissues (66). Nevertheless, aminoglycoside
melanotic dopaminergic neurons in the substantia nigra of the
therapy causes significantly more toxicity in albino animals
brain. A provocative connection between the drug-binding
than in their pigmented counterparts. The administration of
properties of melanin and the etiology of Parkinson’s disease
aminoglycosides can result in permanent vestibular and audi-
came from the observation that heroin contaminated with
tory ototoxicity (7). The cochlear melanin content has been
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) caused
correlated to the pigmentation of the host (65). The ototoxic
a similar neurological disease in drug users, possibly because
effects of aminoglycosides have been shown by electrophysio-
melanin concentrated this compound in substantia nigra neu-
logical and morphological methods (19, 136, 143). However,
rons (46). Chlorpromazine also accumulates in the substantia
no difference in outer hair cell degeneration in the organs of
nigra (73), and the side effects of chlorpromazine and other
Corti was observed between albino and pigmented guinea pigs
phenothiazines include extrapyramidal disorders, such as tar-
exposed to various concentrations of kanamycin (137). Ami-
dive dyskinesia and parkinsonism. In contrast to MPTP, the
noglycosides can also cause functional and morphological
parkinsonian symptoms secondary to phenothiazines are usu-
changes in the retina, particularly when they are administered
ally reversible. The specific retention of other drugs in pig-
by intravitreal injection. Studies with albino and pigmented
mented tissues can damage cells in the skin, eye, and inner ear.
rabbits have shown that ocular pigmentation can partially pro-
These interactions are complex and depend on diverse factors,
tect the retina from damage due to aminoglycosides (30). Al-
such as cysteine content, pH, and ionic interactions (79). As
though it is of unclear significance, the binding of aminoglyco-
described above for synthetic DOPA melanin, Scatchard plot
side by melanin can augment the antibiotic’s inhibitory effects
analysis has revealed the heterogeneity of binding sites on
on collagen synthesis in human fibroblasts in vitro (141).
melanin for single compounds (69, 102, 122, 123). The confor-
The capacity of melanin to bind to aminoglycosides and
mation of the compounds may also influence these interac-
other antibiotics may have important implications when these
tions. Binding is typically reversible, but the retention times
drugs are used in intraocular injections (4, 63). The in vitro
can be protracted. For example, chloroquine can be detected
efficacies of aminoglycosides, tertracyclines, and vancomyin
in the melanin of the eye for a year after receipt of a single
were significantly reduced following incubation with mela-
dose (74), and chloroquine therapy is associated with retinop-
nin (5, 38). In fact, the mixing of 100 g/ml of tobramycin
athies (49) that can occur long after treatment (151). In addi-
with 1,000 g/ml of melanin resulted in an immediate de-
tion to chloroquine, severe retinopathies can occur following
crease in antibiotic activity of 80% (5). The efficacies of
melanin binding by chlorpromazine. Chloroquine also accumu-
fluoroquinolones may also be affected, as these drugs are
lates in dermal melanocytes and hair follicles (79), where it
bound by melanin within the eye (37, 39, 40) and even in
can occasionally cause irreversible hearing loss, tinnitus, and
hair (140). Although melanin can bind to fluoroquinolones,
dizziness (44). Whereas hearing loss due to chloroquine is
penicillins, and cephalosporins, no reduction in antibacterial
thought to be a result of effects on the eighth cranial nerve,
efficacy has been reported after these drugs have been in-
quinine can accumulate in melanin in the stria vascularis of the
cochlea and cause cellular degeneration (74).
Thioureylenes are selectively incorporated into melanin
MELANIN AND THE EFFICACY OF
(71); and certain compounds, such as propylthiouracil, may
ANTIMICROBIAL THERAPY
cause a loss or the depigmentation of hair (70). The carcino-genic effects of polycyclic aromatic hydrocarbons may be com-
Melanin has been called an “an antifungal resistance factor,”
pounded by the presence of melanin, since these compounds
given its ability to reduce the susceptibilities of melanized cells
have a prolonged retention time in pigmented tissues (106).
to antifungal drugs (52). Notably, there is no evidence for the
Herbicides, such as paraquat (which is structurally related to
involvement of melanin in drug efflux pumps or in alterations
1-methyl-4-phenylpyridinium [MPPϩ], the neurotoxic metabo-
in the synthesis of ergosterols or glucans in fungal cell wall/cell
lite of MPTP), avidly bind to melanin and cause parkinsonian
symptoms in experimental animals (3). Cocaine and amphet-
Antifungal susceptibility testing. In vitro susceptibility mea-
amines are known to bind to melanin (12, 119), and testing of
sures the activity of a drug against a microbe, whereas clinical
hair for these compounds is used for medical and legal pur-
resistance is a lack of efficacy of a drug in vivo. Although in
poses. The cytotoxic effects of anthracycline chemotherapeu-
vitro resistance often correlates with clinical treatment failure,
tics (such as doxorubicin and donorubicin) can be inhibited by
in vitro susceptibility does not necessarily predict clinical suc-
melanin. For example, the 50% inhibitory concentration of
cess. Standard MIC broth macrodilution testing by use of the
donorubicin in an in vitro cell-based assay increased from 0.04
M27A protocol for yeasts of the Clinical and Laboratory Stan-
to 0.08 M in the presence of melanin from S. officinalis (121).
dards Institute (CLSI; formerly the National Committee of
TABLE 3. Melanin production protects pathogenic fungi
(124). Melanization was also associated with reduced suscep-
tibility to amphotericin B in melanized versus nonmelanized B. dermatitidis
cells (96). Melanized P. brasiliensis cells were also
more resistant to amphotericin B, fluconazole, ketoconazole,itraconazole, and sulfamethoxazole than nonmelanized cells
(21). However, melanization can also increase susceptibility to
certain drugs. The antipsychotic drug trifluoperazine had
greater fungicidal activity against melanized cryptococcal cells
than it did against nonmelanized cryptococcal cells when ac-tivity was measured by both CFU determination and flow cy-
tometry with propidium iodide staining (135). In contrast,chloroquine, which is highly bound by melanin, had no fungi-
cidal effect on either melanized or nonmelanized C. neofor-mans cells (135). Hence, an association between the capacity of
melanin to bind to a drug and reduced susceptibility to thatdrug by melanized cells is not apparent for all agents. This
a Magnitude indicates the maximal percent increase in protection afforded to
implies the existence of mechanisms other than simple absorp-
the organism by the production of melanin compared to that afforded to cellsdeficient in melanin.
tion by melanin as an explanation for the differences in theactivities of certain classes of drugs against melanized andnonmelanized cells.
Clinical Laboratory Standards) revealed no differences in sus-
The efficacies of antifungals to melanized cells can also
ceptibility between melanized and nonmelanized C. neofor-
be evaluated by a 2,3-bis(2-methoxy-4-nitro-5-sulfophenyl)-5-
mans cells (52, 124). Similarly, no differences in susceptibility
[(phenylamino)carbonyl]-2H-tetrazolium hydroxide (XTT) re-
to antifungals by MIC assays were measured between albino
duction assay (80). Specifically, XTT was used to show that
and pigmented cells of E. dermatitidis (101), H. capsulatum
melanization protected C. neoformans from amphotericin B
(124), or Blastomyces dermatitidis (96). However, the growth of
and caspofungin in biofilms. For example, the metabolic activ-
melanized C. neoformans yeast cells in medium without a phe-
ity of melanized C. neoformans cells in a biofilm exposed to 32
nolic substrate resulted in large defects in the melanin layer of
g/ml of amphotericin B was 40%, whereas it was 20% for
the parent cells after budding and an absence of melanin in the
daughter cells (89). Hence, even if melanized cells are used
Impact of melanin binding on antifungal drugs. The finding
initially, the daughter cells lack melanin, and the CLSI meth-
that melanin can bind to amphotericin B and caspofungin, in
odology does not compare the susceptibilities of melanized
combination with observations of the microstructure of mela-
and nonmelanized cells. Incorporation of phenolic substrates
nin in C. neoformans, suggests a potential explanation for the
into the testing medium for a microdilution or a macrodilution
difficulty in eradicating C. neoformans with these drugs. Mel-
assay has not been possible because these either precipitate or
anization of C. neoformans yeast cells occurs in vivo (93), and
autopolymerize into melanin (124). Hence, the CLSI protocol
the amount of melanin produced increases with time after
is not suitable for distinguishing differences in susceptibility
infection (31, 95). Although amphotericin B is fungicidal to
between melanized and nonmelanized cells.
nonmelanized C. neoformans cells in vitro, amphotericin B
In contrast to the broth dilution method, time-kill studies
therapy often fails to eradicate the fungus from patients (100,
with CFU determinations have revealed differences in the sus-
152). Given the relative resistance of melanized cells to am-
ceptibilities of melanized and albino cells to certain antifungal
photericin B, the efficacy of this drug may be due to its activity
drugs (52, 124, 125) (Table 3). A major difference between
against nonmelanized buds and to its facilitation of tissue
these assays is that in time-kill studies the microbes are incu-
clearance by host defenses through its powerful immunomodu-
bated with the antimicrobial drugs for hours rather than days.
lating effects. Caspofungin is active in vitro against nonmela-
Thus, the melanin layer of the fungus remains largely intact. C.
nized cells (36), but it is ineffective against experimental infec-
neoformans is significantly less susceptible to amphotericin B
tions with C. neoformans (1). The inefficacy of caspofungin for
when the fungus is grown in the presence of L-dopa (133). This
C. neoformans in animal studies cannot be explained by its
result was recently confirmed by time-kill studies with ampho-
inability to inhibit either 1-3--D- or 1-6--D-glucan synthase
tericin and caspofungin, which revealed that the activities of
(32). Since acapsular strains of C. neoformans are also resistant
these drugs against melanized cells were reduced by 55 and
to caspofungin, the large polysaccharide capsules that can oc-
7%, respectively, relative to their activities against nonmela-
cur in vivo are not inhibiting the drug from engaging the
nized cells (52, 124). In contrast, no differences in activity
fungus. Hence, for C. neoformans, the clinical resistance to
against melanized and nonmelanized C. neoformans cells was
caspofungin could be attributed in part to in vivo yeast cell
observed for voriconazole, fluconazole, itraconazole, or flucy-
melanization. Caspofungin is clinically effective against
tosine (124, 125). However, melanized C. neoformans strains
Aspergillus spp., a group of fungi that can produce melanin.
exhibited reduced susceptibilities to higher concentrations of
However, its efficacy may be due to the fact that hyphae, the
tissue-invasive form of this fungus, are not melanized (146).
Time-kill assays similarly revealed that melanized H. capsu-
Dematiaceous fungi are darkly pigmented molds that con-
latum yeast cells were less susceptible to amphotericin B and
stitutively produce melanin during infection and are extremely
caspofungin than nonmelanized H. capsulatum yeast cells
difficult to treat with antifungal drugs (13). Although the clin-
ical relevance is not clear, antifungal susceptibility testing for
fact that voriconazole at 0.125 to 0.5 mg/liter can inhibit
filamentous fungi has recently been standardized (86). Ampho-
conidiation in diverse Aspergillus spp., resulting in white colo-
tericin B has good activity against most clinically important
nies (126). Ravuconazole, which is structurally similar to vori-
dematiaceous fungi in vitro, but clinical resistance is not un-
conazole, had similar effects only against Aspergillus fumigatus
common. Scedosporium prolificans and Scopulariopsis brumptii
and Aspergillus flavus. It is possible that the inhibition of mel-
are consistently resistant to amphotericin B in vitro; and occa-
anin formation in vivo may contribute to the therapeutic
sional resistance to this drug is reported in several other
potencies of these triazoles by increasing the susceptibility to
species, including Chaetomium spp., Curvularia spp., Phiale-
host defense mechanisms. The possibility that certain antifun-
monium spp., and Exophiala spp. (81). Echinocandins are not
gal agents are less effective against melanotic molds should
clinically useful against these fungi. The inefficacies of the
especially be considered when clinicians make choices for em-
echinocandins against these fungi and the relative resistance of
pirical therapy in patients with presumed mycotic diseases.
these fungi to amphotericin B may be associated with thedense production of melanin in these fungi. The broadest in
ACKNOWLEDGMENTS
vitro activity against dematiaceous fungi is achieved with azoles
J.D.N. and A.C. are supported in part by NIH grant AI52733.
(81). In this regard, we note that azoles are not bound by
The electron microscopy images in Fig. 2 are courtesy of Helene
Impact of melanin binding on antibacterial agents. The role REFERENCES
of melanin in the protection of bacteria from antimicrobial
1. Abruzzo, G. K., A. M. Flattery, C. J. Gill, L. Kong, J. G. Smith, V. B.
drugs is largely unexplored. Recently, an Escherichia coli strain
Pikounis, J. M. Balkovec, A. F. Bouffard, J. F. Dropinski, H. Rosen, H.
expressing a recombinant plasmid containing a tyrosinase gene
Kropp, and K. Bartizal. 1997. Evaluation of the echinocandin antifungal
was constructed, and the E. coli strain produced melanin in
MK-0991 (L-743,872): efficacies in mouse models of disseminated aspergil-losis, candidiasis, and cryptococcosis. Antimicrob. Agents Chemother.
medium supplemented with tyrosine (72). In contrast, Pseudo-monas aeruginosa melanin does not appear to serve a protec-
2. Alviano, D. S., A. J. Franzen, L. R. Travassos, C. Holandino, S. Rozental,
tive role against antibacterial agents in vitro (111, 112). How-
R. Ejzemberg, C. S. Alviano, and M. L. Rodrigues. 2004. Melanin from Fonsecaea pedrosoi induces production of human antifungal antibodies and
ever, melanin in Bacillus thuringiensis can protect the bacteria
enhances the antimicrobial efficacy of phagocytes. Infect. Immun. 72:229–
against pesticides (98). Since little is known about the location
3. Barbeau, A., L. Dallaire, N. T. Buu, J. Poirier, and E. Rucinska. 1985.
of the pigment in bacteria that produce melanin, the protective
Comparative behavioral, biochemical and pigmentary effects of MPTP,
effects may be limited. For example, if melanin is intracellular,
MPPϩ and paraquat in Rana pipiens. Life Sci. 37:1529–1538.
antibiotics such as penicillins that target the cell wall would not
4. Barza, M. 1978. Factors affecting the intraocular penetration of antibiotics.
The influence of route, inflammation, animal species and tissue pigmenta-
be expected to interact with the polymer before they reach
tion. Scand. J. Infect. Dis. Suppl, p. 151–159.
5. Barza, M., J. Baum, and A. Kane. 1976. Inhibition of antibiotic activity in
vitro by synthetic melanin. Antimicrob. Agents Chemother. 10:569–570.
6. Bathory, G., T. Szuts, and K. Magyar. 1987. Studies on the melanin affinity CONCLUSIONS
of selegiline (deprenyl) and other amphetamine derivatives. Pol. J. Phar- macol. Pharm. 39:195–201.
7. Black, F. O., S. Pesznecker, and V. Stallings. 2004. Permanent gentamicin
Melanin production provides survival advantages to myriad
vestibulotoxicity. Otol. Neurotol. 25:559–569.
microbes in the environment and during infection of diverse
8. Blasi, E., R. Barluzzi, R. Mazzolla, B. Tancini, S. Saleppico, M. Puliti, L.
hosts. There is conclusive evidence that many types of drugs,
Pitzurra, and F. Bistoni. 1995. Role of nitric oxide and melanogenesis in the accomplishment of anticryptococcal activity by the BV-2 microglial cell
including antimicrobial drugs, bind to melanin. In particular,
line. J. Neuroimmunol. 58:111–116.
the melanization of certain fungi is associated with reduced
9. Bloomfield, B. J., and M. Alexander. 1967. Melanins and resistance of fungi
susceptibilities to polyene and echinocandin-type drugs in
to lysis. J. Bacteriol. 93:1276–1280.
10. Bocca, A. L., P. P. Brito, F. Figueiredo, and C. E. Tosta. 2006. Inhibition of
vitro. In contrast, melanization has not been associated with
nitric oxide production by macrophages in chromoblastomycosis: a role for
reduced susceptibilities to azole-type drugs, except at high con-
Fonsecaea pedrosoi melanin. Mycopathologia 161:195–203.
11. Borel, M., D. Lafarge, M. F. Moreau, M. Bayle, L. Audin, N. Moins, and
centrations. Current standard methods for antifungal drug sus-
J. C. Madelmont. 2005. High resolution magic angle spinning NMR spec-
ceptibility testing are not adequate to discern a melanin effect
troscopy used to investigate the ability of drugs to bind to synthetic melanin.
on antifungal drug activity, and measurement of this effect
Pigment Cell Res. 18:49–54.
12. Borges, C. R., J. C. Roberts, D. G. Wilkins, and D. E. Rollins. 2003.
requires the use of time-kill assays or an assessment of meta-
Cocaine, benzoylecgonine, amphetamine, and N-acetylamphetamine bind-
bolic activity. In vivo data demonstrate that compounds that
ing to melanin subtypes. J. Anal. Toxicol. 27:125–134.
inhibit melanization can reduce the virulence of C. neoformans
13. Brandt, M. E., and D. W. Warnock. 2003. Epidemiology, clinical manifes-
tations, and therapy of infections caused by dematiaceous fungi. J. Che-
and other fungi. The administration of monoclonal antibodies
mother. 15:36–47.
to melanin or glyphosate (which inhibits the melanization of C.
14. Bridelli, M. G., A. Ciati, and P. R. Crippa. 2006. Binding of chemicals to
melanins re-examined: adsorption of some drugs to the surface of melanin
neoformans) prolongs the survival of mice lethally infected with
particles. Biophys. Chem. 119:137–145. C. neoformans (92, 110). Similarly, several studies have dem-
15. Bull, A. T. 1970. Inhibition of polysaccharases by melanin: enzyme inhibi-
onstrated that melanin-deficient E. dermatitidis strains are less
tion in relation to mycolysis. Arch. Biochem. Biophys. 137:345–356.
16. Buszman, E., and R. Rozanska. 2003. Interaction of quinidine, disopyr-
virulent than melanized strains (22–24, 33). The development
amide and metoprolol with melanin in vitro in relation to drug-induced
of drugs that interfere with melanin polymerization or rear-
ocular toxicity. Pharmazie 58:507–511.
rangement may be useful therapeutic compounds for the treat-
17. Butler, M. J., and A. W. Day. 1998. Fungal melanins: a review. Can. J.
Microbiol. 44:1115–1136.
ment of these melanotic fungi and other pathogens that pro-
18. Castanet, J., and J. P. Ortonne. 1997. Hair melanin and hair color. EXS
duce melanin (91). Also, it is possible that the use of agents
78:209–225.
that inhibit melanization may render melanotic fungi suscep-
Conlee, J. W., M. L. Bennett, and D. J. Creel. 1995. Differential effects of gentamicin on the distribution of cochlear function in albino and pigmented
tible to drugs that bind to melanin. An interesting finding is the
guinea pigs. Acta Otolaryngol. 115:367–374.
20. Cunha, M. M., A. J. Franzen, D. S. Alviano, E. Zanardi, C. S. Alviano, W.
mycelium: melanins of phytopathogenic fungi. Annu. Rev. Phytopathol. De Souza, and S. Rozental. 2005. Inhibition of melanin synthesis pathway 37:447–471.
by tricyclazole increases susceptibility of Fonsecaea pedrosoi against mouse
46. Herrero, M. T., E. C. Hirsch, A. Kastner, M. Ruberg, M. R. Luquin, J.
macrophages. Microsc. Res. Tech. 68:377–384. Laguna, F. Javoy-Agid, J. A. Obeso, and Y. Agid. 1993. Does neuromelanin
21. da Silva, M. B., A. F. Marques, J. D. Nosanchuk, A. Casadevall, L. R.
contribute to the vulnerability of catecholaminergic neurons in monkeys
Travassos, and C. P. Taborda. 2006. Melanin in the dimorphic fungal
intoxicated with MPTP? Neuroscience 56:499–511.
pathogen Paracoccidioides brasiliensis: effects on phagocytosis, intracellular
47. Hill, H. Z. 1991. Melanins in the photobiology of skin cancer and the
resistance and drug susceptibility. Microbes Infect. 8:197–205.
radiobiology of melanomas, p. 31–53. In S. H. Wilson (ed.), Cancer biology
22. Dixon, D. M., J. Migliozzi, C. R. Cooper, Jr., O. Solis, B. Breslin, and P. J.
and biosynthesis. Telford Press, Caldwell, N.J. Szaniszlo. 1992. Melanized and non-melanized multicellular form mutants
48. Hill, H. Z. 1992. The function of melanin or six blind people examine an
of Wangiella dermatitidis in mice: mortality and histopathology studies.
elephant. Bioessays 14:49–56.
Mycoses 35:17–21.
49. Hobbs, H. E., A. Sorsby, and A. Freedman. 1959. Retinopathy following
23. Dixon, D. M., A. Polak, and G. W. Conner. 1989. MelϪ mutants of Wangiella
chloroquine therapy. Lancet ii:478–480. dermatitidis in mice: evaluation of multiple mouse and fungal strains.
50. Huffnagle, G. B., G. H. Chen, J. L. Curtis, R. A. McDonald, R. M. Strieter,
J. Med. Vet. Mycol. 27:335–341. and G. B. Toews. 1995. Down-regulation of the afferent phase of T cell-
24. Dixon, D. M., A. Polak, and P. J. Szaniszlo. 1987. Pathogenicity and viru-
mediated pulmonary inflammation and immunity by a high melanin-pro-
lence of wild-type and melanin-deficient Wangiella dermatitidis. J. Med. Vet.
ducing strain of Cryptococcus neoformans. J. Immunol. 155:3507–3516.
Mycol. 25:97–106.
51. Icenhour, C. R., T. J. Kottom, and A. H. Limper. 2006. Pneumocystis
25. Dixon, D. M., P. J. Szaniszlo, and A. Polak. 1991. Dihydroxynaphthalene
melanins confer enhanced organism viability. Eukaryot. Cell 5:916–923.
(DHN) melanin and its relationship with virulence in the early stages of
52. Ikeda, R., T. Sugita, E. S. Jacobson, and T. Shinoda. 2003. Effects of
phaeohyphomycosis, p. 297–318. In G. Cole and H. Hoch (ed.), The fungal
melanin upon susceptibility of Cryptococcus to antifungals. Microbiol. Im-
spore and disease initiation in plants and animals. Plenum Press, New York,
munol. 47:271–277.
53. Ings, R. 1984. The melanin binding of drugs and its implications. Drug
26. Doering, T. L., J. D. Nosanchuk, W. K. Roberts, and A. Casadevall. 1999.
Metab. Rev. 15:1183–1212.
Melanin as a potential cryptococcal defence against microbicidal proteins.
54. Jacobson, E. S. 2000. Pathogenic roles for fungal melanins. Clin. Microbiol.
Med. Mycol. 37:175–181.
Rev. 13:708–717.
27. Eisenman, H. C., J. D. Nosanchuk, J. B. Webber, R. J. Emerson, T. A.
55. Jacobson, E. S., and H. S. Emery. 1991. Catecholamine uptake, melaniza- Camesano, and A. Casadevall. 2005. Microstructure of cell wall-associated
tion, and oxygen toxicity in Cryptococcus neoformans. J. Bacteriol. 173:401–
melanin in the human pathogenic fungus Cryptococcus neoformans. Bio-
chemistry 44:3683–3693.
56. Jacobson, E. S., and J. D. Hong. 1997. Redox buffering by melanin and
28. Emery, H. S., C. P. Shelburne, J. P. Bowman, P. G. Fallon, C. A. Schulz, and
Fe(II) in Cryptococcus neoformans. J. Bacteriol. 179:5340–5346. E. S. Jacobson. 1994. Genetic study of oxygen resistance and melanization
57. Jacobson, E. S., and R. Ikeda. 2005. Effect of melanization upon porosity of
in Cryptococcus neoformans. Infect. Immun. 62:5694–5697.
the cryptococcal cell wall. Med. Mycol. 43:327–333.
29. Enochs, W. S., M. J. Nilges, and H. M. Swartz. 1993. A standardized test for
58. Jacobson, E. S., N. D. Jenkins, and J. M. Todd. 1994. Relationship between
the identification and characterization of melanins using electron paramag-
superoxide dismutase and melanin in a pathogenic fungus. Infect. Immun.
netic (EPR) spectroscopy. Pigment Cell Res. 6:91–99. 62:4085–4086.
30. Eves, P., L. Smith-Thomas, S. Hedley, M. Wagner, C. Balafa, and S.
59. Jacobson, E. S., and S. B. Tinnell. 1993. Antioxidant function of fungal MacNeil. 1999. A comparative study of the effect of pigment on drug
melanin. J. Bacteriol. 175:7102–7104.
toxicity in human choroidal melanocytes and retinal pigment epithelial
60. Jahn, B., A. Koch, A. Schmidt, G. Wanner, H. Gehringer, S. Bhakdi, and
cells. Pigment Cell Res. 12:22–35. A. A. Brakhage. 1997. Isolation and characterization of a pigmentless-
31. Feldmesser, M., Y. Kress, and A. Casadevall. 2001. Dynamic changes in the
conidium mutant of Aspergillus fumigatus with altered conidial surface and
morphology of Cryptococcus neoformans during murine pulmonary infec-
reduced virulence. Infect. Immun. 65:5110–5117.
tion. Microbiology 147:2355–2365.
61. Joseph, R. E., Jr., W. J. Tsai, L. I. Tsao, T. P. Su, and E. J. Cone. 1997. In
32. Feldmesser, M., Y. Kress, A. Mednick, and A. Casadevall. 2000. The effect
vitro characterization of cocaine binding sites in human hair. J. Pharmacol.
of the echinocandin analogue caspofungin on cell wall glucan synthesis by
Exp. Ther. 282:1228–1241. Cryptococcus neoformans. J. Infect. Dis. 182:1791–1795.
62. Kaliszan, R., A. Kaliszan, and I. W. Wainer. 1993. Prediction of drug
33. Feng, B., X. Wang, M. Hauser, S. Kaufmann, S. Jentsch, G. Haase, J. M.
binding to melanin using a melanin-based high-performance liquid chro-
Becker, and P. J. Szaniszlo. 2001. Molecular cloning and characterization
matographic stationary phase and chemometric analysis of the chromato-
of WdPKS1, a gene involved in dihydroxynaphthalene melanin biosynthesis
graphic data. J. Chromatogr. 615:281–288.
and virulence in Wangiella (Exophiala) dermatitidis. Infect. Immun. 69:
63. Kane, A., M. Barza, and J. Baum. 1981. Intravitreal injection of gentamicin
in rabbits. Effect of inflammation and pigmentation on half-life and ocular
34. Fiore, G., A. Poli, A. Di Cosmo, M. d’Ischia, and A. Palumbo. 2004.
distribution. Investig. Ophthalmol. Vis. Sci. 20:593–597.
Dopamine in the ink defense system of Sepia officinalis: biosynthesis, ve-
64. Kawamura, C., T. Tsujimoto, and T. Tsuge. 1999. Targeted disruption of a
sicular compartmentation in mature ink gland cells, nitric oxide (NO)/
melanin biosynthesis gene affects conidial development and UV tolerance
cGMP-induced depletion and fate in secreted ink. Biochem. J. 378:785–791.
in the Japanese pear pathotype of Alternaria alternata. Mol. Plant-Microbe
35. Fogarty, R. V., and J. M. Tobin. 1996. Fungal melanins and their interac-
Interact. 12:59–63.
tions with metals. Enzyme Microb. Technol. 19:311–317.
65. Keithley, E. M., A. F. Ryan, and M. L. Feldman. 1992. Cochlear degener-
36. Franzot, S. P., and A. Casadevall. 1997. Pneumocandin L-743,872 enhances
ation in aged rats of four strains. Hear. Res. 59:171–178.
the activities of amphotericin B and fluconazole against Cryptococcus neo-
66. Kumana, C. R., and K. Y. Yuen. 1994. Parenteral aminoglycoside therapy. formans in vitro. Antimicrob. Agents Chemother. 41:331–336.
Selection, administration and monitoring. Drugs 47:902–913.
37. Fukuda, M., Y. Morita, K. Sasaki, and Y. Yamamoto. 2000. Studies on the
67. Kuo, M. J., and M. Alexander. 1967. Inhibition of the lysis of fungi by
binding mechanism of fluoroquinolones to melanin. J. Infect. Chemother.
melanins. J. Bacteriol. 94:624–629. 6:72–76.
68. Lamy-Freund, M. T., S. Schreier, R. M. Peitzsch, and W. F. Reed. 1991.
38. Fukuda, M., and K. Sasaki. 1990. Changes in the antibacterial activity of
Characterization and time dependence of amphotericin B: deoxycholate
melanin-bound drugs. Ophthalmic Res. 22:123–127.
aggregation by quasielastic light scattering. J. Pharm. Sci. 80:262–266.
39. Fukuda, M., and K. Sasaki. 1995. Differences between albino and pig-
69. Larsson, B., and H. Tjalve. 1979. Studies on the mechanism of drug-binding
mented rabbit eyes in the intraocular pharmacokinetics of sparfloxacin.
to melanin. Biochem. Pharmacol. 28:1181–1187.
Drugs 49:314–316.
70. Larsson, B. S. 1993. Interaction between chemicals and melanin. Pigment
40. Fukuda, M., and K. Sasaki. 1994. Different iris coloration and uptake of a
Cell Res. 6:127–133.
fluoroquinolone agent into the iris ciliary body of rabbit eyes. Ophthalmic
71. Larsson, B. S. 1991. Melanin-affinic thioureas as selective melanoma
Res. 26:137–140.
seekers. Melanoma Res. 1:85–90.
41. Gan, E. V., H. F. Haberman, and I. A. Menon. 1976. Electron transfer
72. Lin, W. P., H. L. Lai, Y. L. Liu, Y. M. Chiung, C. Y. Shiau, J. M. Han, C. M.
properties of melanin. Arch. Biochem. Biophys. 173:666–672. Yang, and Y. T. Liu. 2005. Effect of melanin produced by a recombinant
42. Garcia-Rivera, J., and A. Casadevall. 2001. Melanization of Cryptococcus Escherichia coli on antibacterial activity of antibiotics. J. Microbiol. Immu-
neoformans reduces its susceptibility to the antimicrobial effects of silver
nol. Infect. 38:320–326.
nitrate. Med. Mycol. 39:353–357.
73. Lindquist, N. G. 1972. Accumulation in vitro of 35S-chlorpromazine in the
43. Gautam, L., K. S. Scott, and M. D. Cole. 2005. Amphetamine binding to
neuromelanin of human substantia nigra and locus coeruleus. Arch. Int.
synthetic melanin and Scatchard analysis of binding data. J. Anal. Toxicol.
Pharmacodyn. Ther. 200:190–195. 29:339–344.
74. Lindquist, N. G. 1973. Accumulation of drugs on melanin. Acta Radiol.
44. Hart, C. W., and R. F. Naunton. 1964. The ototoxicity of chloroquine
Diagn. (Stockholm) 325:1–92.
phosphate. Arch. Otolaryngol. 80:407–412.
75. Liu, G. Y., A. Essex, J. T. Buchanan, V. Datta, H. M. Hoffman, J. F. Bastian,
45. Henson, J. M., M. J. Butler, and A. W. Day. 1999. The dark side of the J. Fierer, and V. Nizet. 2005. Staphylococcus aureus golden pigment impairs
neutrophil killing and promotes virulence through its antioxidant activity. J.
102. Potsch, L., G. Skopp, and G. Rippin. 1997. A comparison of 3H-cocaine
Exp. Med. 202:209–215.
binding on melanin granules and human hair in vitro. Int. J. Legal Med.
76. Liu, L., R. P. Tewari, and P. R. Williamson. 1999. Laccase protects 110:55–62. Cryptococcus neoformans from antifungal activity of alveolar macrophages.
103. Prota, G. 1992. Melanins and melanogenesis. Academic Press, Inc., San
Infect. Immun. 67:6034–6039.
77. Lukiewicz, S., K. Reszka, and Z. Matusak. 1980. Simultaneous electo-
104. Richman, A., and F. C. Kafatos. 1996. Immunity to eukaryotic parasites in
chemical-electron spin resonance (SEESR) studies on natural and synthetic
vector insects. Curr. Opin. Immunol. 8:14–19.
melanins. Bioelectrochem. Bioenerg. 7:153–165.
105. Rizzo, D., R. Blanchette, and M. Palmer. 1992. Biosorption of metal ions by
78. Marmaras, V. J., N. D. Charalambidis, and C. G. Zervas. 1996. Immune Armillaria rhizomorphus. Can. J. Bot. Rev. 70:1515–1520.
response in insects: the role of phenoloxidase in defense reactions in rela-
106. Roberto, A., B. S. Larsson, and H. Tjalve. 1996. Uptake of 7,12-dimethyl-
tion to melanization and sclerotization. Arch. Insect Biochem. Physiol.
benz(a)anthracene and benzo(a)pyrene in melanin-containing tissues. 31:119–133.
Pharmacol. Toxicol. 79:92–99.
79. Mars, U., and B. S. Larsson. 1999. Pheomelanin as a binding site for drugs
107. Romero-Martinez, R., M. Wheeler, A. Guerrero-Plata, G. Rico, and H.
and chemicals. Pigment Cell Res. 12:266–274. Torres-Guerrero. 2000. Biosynthesis and functions of melanin in Sporothrix
80. Martinez, L. R., and A. Casadevall. 2006. Susceptibility of Cryptococcus schenckii. Infect. Immun. 68:3696–3703. neoformans biofilms to antifungal agents in vitro. Antimicrob. Agents Che-
108. Rosas, A. L., and A. Casadevall. 1997. Melanization affects susceptibility
mother. 50:1021–1033.
of Cryptococcus neoformans to heat and cold. FEMS Microbiol. Lett. 153:
81. McGinnis, M. R., and L. Pasarell. 1998. In vitro testing of susceptibilities of
filamentous ascomycetes to voriconazole, itraconazole, and amphotericin B,
109. Rosas, A. L., and A. Casadevall. 2001. Melanization decreases the sus-
with consideration of phylogenetic implications. J. Clin. Microbiol. 36:
ceptibility of Cryptococcus neoformans to enzymatic degradation. Myco-
pathologia 151:53–56.
82. Mednick, A. J., J. D. Nosanchuk, and A. Casadevall. 2005. Melanization
110. Rosas, A. L., J. D. Nosanchuk, and A. Casadevall. 2001. Passive immuni-
of Cryptococcus neoformans affects lung inflammatory responses during
zation with melanin-binding monoclonal antibodies prolongs survival of
cryptococcal infection. Infect. Immun. 73:2012–2019.
mice with lethal Cryptococcus neoformans infection. Infect. Immun. 69:
83. Mironenko, N. V., I. A. Alekhina, N. N. Zhdanova, and S. A. Bulat. 2000.
Intraspecific variation in gamma-radiation resistance and genomic structure
111. Rozhavin, M. A. 1978. Effect of Pseudomonas aeruginosa melanin on anti-
in the filamentous fungus Alternaria alternata: a case study of strains inhab-
biotic activity. Antibiotiki 23:718–720.
iting Chernobyl Reactor No. 4. Ecotoxicol. Environ. Safety 45:177–187.
112. Rozhavin, M. A., and V. V. Sologub. 1979. Comparison of the sensitivity of
84. Mylonakis, E., F. M. Ausubel, J. R. Perfect, J. Heitman, and S. B. Calderwood. Pseudomonas aeruginosa cultures that synthesize melanin and other pig-
2002. Killing of Caenorhabditis elegans by Cryptococcus neoformans as a model
ments to 12 antibiotics and 5-nitro-8-quinolinol. Antibiotiki 24:921–922.
of yeast pathogenesis. Proc. Natl. Acad. Sci. USA 99:15675–15680.
113. Saleh, Y. G., M. S. Mayo, and D. G. Ahearn. 1988. Resistance of some
85. Nappi, A. J., and B. M. Christensen. 2005. Melanogenesis and associated
common fungi to gamma irradiation. Appl. Environ. Microbiol. 54:2134–
cytotoxic reactions: applications to insect innate immunity. Insect Biochem.
Mol. Biol. 35:443–459.
114. Sanchez-Ferrer, A., J. N. Rodriguez-Lopez, F. Garcia-Canovas, and F.
86. National Committee for Clinical Laboratory Standards. 2002. Reference Garcia-Carmona. 1995. Tyrosinase: a comprehensive review of its mecha-
method for broth dilution antifungal susceptibility testing of conidium-
nism. Biochim. Biophys. Acta 1247:1–11.
forming filamentous fungi. Approved standard M38-A. National Commit-
115. Schnitzler, N., H. Peltroche-Llacsahuanga, N. Bestier, J. Zundorf, R.
tee for Clinical Laboratory Standards, Wayne, Pa. Lutticken, and G. Haase. 1999. Effect of melanin and carotenoids of
87. Nicholaus, R., M. Piatelli, and E. Fattorusso. 1964. The structure of mel- Exophiala (Wangiella) dermatitidis on phagocytosis, oxidative burst, and
anins and melanogenesis. IV. On some natural melanins. Tetrahedron
killing by human neutrophils. Infect. Immun. 67:94–101. 20:1163–1172.
116. Sichel, G., C. Corsaro, M. Scalia, A. J. Di Bilio, and R. P. Bonomo. 1991. In
88. Nosanchuk, J., and A. Casadevall. 1997. Cellular charge of Cryptococcus
vitro scavenger activity of some flavonoids and melanins against O2Ϫ · . neoformans: contributions from the capsular polysaccharide, melanin, and
Free Radic. Biol. Med. 11:1–8.
monoclonal antibody binding. Infect. Immun. 65:1836–1841.
117. Steenbergen, J. N., and A. Casadevall. 2003. The origin and maintenance of
89. Nosanchuk, J. D., and A. Casadevall. 2003. Budding of melanized Crypto-
virulence for the human pathogenic fungus Cryptococcus neoformans. Mi-
coccus neoformans in the presence or absence of L-dopa. Microbiology
crobes Infect. 5:667–675. 149:1945–1951.
118. Steenbergen, J. N., H. A. Shuman, and A. Casadevall. 2001. Cryptococcus
90. Nosanchuk, J. D., and A. Casadevall. 2003. The contribution of melanin to neoformans interactions with amoebae suggest an explanation for its viru-
microbial pathogenesis. Cell. Microbiol. 5:203–223.
lence and intracellular pathogenic strategy in macrophages. Proc. Natl.
91. Nosanchuk, J. D., A. Casadevall, and R. Ovalle. January 2003. Method for
Acad. Sci. USA 98:15245–15250.
inhibiting melanogenesis and uses thereof. U.S. patent 6,509,325.
119. Stout, P. R., and J. A. Ruth. 1999. Deposition of [3H]cocaine, [3H]nicotine,
92. Nosanchuk, J. D., R. Ovalle, and A. Casadevall. 2001. Glyphosate inhibits
and [3H]flunitrazepam in mouse hair melanosomes after systemic admin-
melanization of Cryptococcus neoformans and prolongs survival of mice
istration. Drug Metab. Dispos. 27:731–735.
after systemic infection. J. Infect. Dis. 183:1093–1099.
120. Surazynski, A., J. Palka, D. Wrzesniok, E. Buszman, and P. Kaczmarczyk.
93. Nosanchuk, J. D., A. L. Rosas, S. C. Lee, and A. Casadevall. 2000. Mela-
2001. Melanin potentiates daunorubicin-induced inhibition of collagen bio-
nisation of Cryptococcus neoformans in human brain tissue. Lancet 355:
synthesis in human skin fibroblasts. Eur. J. Pharmacol. 419:139–145.
121. Svensson, S. P., S. Lindgren, W. Powell, and H. Green. 2003. Melanin
94. Nosanchuk, J. D., J. Rudolph, A. L. Rosas, and A. Casadevall. 1999. Evi-
inhibits cytotoxic effects of doxorubicin and daunorubicin in MOLT 4 cells.
dence that Cryptococcus neoformans is melanized in pigeon excreta: impli-
Pigment Cell Res. 16:351–354.
cations for pathogenesis. Infect. Immun. 67:5477–5479.
122. Tjalve, H., M. Nilsson, and B. Larsson. 1981. Binding of 14C-spermidine to
95. Nosanchuk, J. D., P. Valadon, M. Feldmesser, and A. Casadevall. 1999.
melanin in vivo and in vitro. Acta Physiol. Scand. 112:209–214.
Melanization of Cryptococcus neoformans in murine infection. Mol. Cell.
123. Tjalve, H., M. Nilsson, and B. Larsson. 1981. Studies on the binding of
Biol. 19:745–750.
chlorpromazine and chloroquine to melanin in vivo. Biochem. Pharmacol.
96. Nosanchuk, J. D., D. van Duin, P. Mandal, P. Aisen, A. M. Legendre, and 30:1845–1847. A. Casadevall. 2004. Blastomyces dermatitidis produces melanin in vitro and
124. Van Duin, D., A. Casadevall, and J. D. Nosanchuk. 2002. Melanization of
during infection. FEMS Microbiol. Lett. 239:187–193. Cryptococcus neoformans and Histoplasma capsulatum reduces their suscep-
97. Nyhus, K. J., A. T. Wilborn, and E. S. Jacobson. 1997. Ferric iron reduction
tibility to amphotericin B and caspofungin. Antimicrob. Agents Chemother.
by Cryptococcus neoformans. Infect. Immun. 65:434–438. 46:3394–3400.
98. Patel, K., J. Wyman, K. Patel, and B. Burden. 1996. A mutant of Bacillus
125. Van Duin, D., W. Cleare, O. Zaragoza, A. Casadevall, and J. D. Nosanchuk. thuringiensis producing a dark-brown pigment with increased UV resistance
2004. Effects of voriconazole on Cryptococcus neoformans. Antimicrob.
and insecticidal activity. J. Invertebr. Pathol. 67:120–124.
Agents Chemother. 48:2014–2020.
99. Peltroche-Llacsahuanga, H., N. Schnitzler, S. Jentsch, A. Platz, S. De Hoog,
126. Varanasi, N. L., I. Baskaran, G. J. Alangaden, P. H. Chandrasekar, and K. G. Schweizer, and G. Haase. 2003. Analyses of phagocytosis, evoked E. K. Manavathu. 2004. Novel effect of voriconazole on conidiation of
oxidative burst, and killing of black yeasts by human neutrophils: a tool for
Aspergillus species. Int. J. Antimicrob. Agents 23:72–79.
estimating their pathogenicity? Med. Mycol. 41:7–14.
127. Vasilevskaya, A., N. M. Zhdanova, and V. D. Pokhodenko. 1970. Character
100. Perfect, J. R., K. A. Marr, T. J. Walsh, R. N. Greenberg, B. DuPont, J. de
of survival of some gamma irradiated species of dark-colored Hyphomycetes. la Torre-Cisneros, G. Just-Nubling, H. T. Schlamm, I. Lutsar, A. Espinel-
Mikrobiol. Zh. 33:438–441. Ingroff, and E. Johnson. 2003. Voriconazole treatment for less-common,
128. Wakamatsu, K., and S. Ito. 2002. Advanced chemical methods in melanin
emerging, or refractory fungal infections. Clin. Infect. Dis. 36:1122–1131.
determination. Pigment Cell Res. 15:174–183.
101. Polak, A., and D. M. Dixon. 1989. Loss of melanin in Wangiella dermatitidis
129. Wakamatsu, K., S. Ito, and J. L. Rees. 2002. The usefulness of 4-amino-3-
does not result in greater susceptibility to antifungal agents. Antimicrob.
hydroxyphenylalanine as a specific marker of pheomelanin. Pigment Cell
Agents Chemother. 33:1639–1640.
Res. 15:225–232.
130. Wang, Y., P. Aisen, and A. Casadevall. 1995. Cryptococcus neoformans
tiates kanamycin-induced inhibition of collagen biosynthesis in human skin
melanin and virulence: mechanism of action. Infect. Immun. 63:3131–3136.
fibroblasts. Pharmazie 60:439–443.
131. Wang, Y., P. Aisen, and A. Casadevall. 1996. Melanin, melanin “ghosts,”
143. Wu, W. J., S. H. Sha, J. D. McLaren, K. Kawamoto, Y. Raphael, and J.
and melanin composition in Cryptococcus neoformans. Infect. Immun. 64: Schacht. 2001. Aminoglycoside ototoxicity in adult CBA, C57BL and
BALB mice and the Sprague-Dawley rat. Hear. Res. 158:165–178.
132. Wang, Y., and A. Casadevall. 1994. Decreased susceptibility of melanized
144. Yabuuchi, E., and A. Ohyama. 1972. Characterization of pyomelanin-pro- Cryptoccocus neoformans to the fungicidal effects of ultraviolet light. Appl.
ducing strains of Pseudomonas aeruginosa. Int. J. Syst. Bacteriol. 22:53–64.
Environ. Microbiol. 60:3864–3866.
145. Yang, Z., R. C. Pascon, A. Alspaugh, G. M. Cox, and J. H. McCusker. 2002.
133. Wang, Y., and A. Casadevall. 1994. Growth of Cryptococcus neoformans in
Molecular and genetic analysis of the Cryptococcus neoformans MET3 gene
presence of L-dopa decreases its susceptibility to amphotericin B. Antimi-
and a met3 mutant. Microbiology 148:2617–2625.
crob. Agents Chemother. 38:2648–2650.
146. Youngchim, S., R. Morris-Jones, R. J. Hay, and A. J. Hamilton. 2004.
134. Wang, Y., and A. Casadevall. 1994. Susceptibility of melanized and non-
Production of melanin by Aspergillus fumigatus. J. Med. Microbiol. 53:175–
melanized Cryptococcus neoformans to nitrogen- and oxygen-derived
oxidants. Infect. Immun. 62:3004–3007.
147. Zecca, L., D. Tampellini, A. Gatti, R. Crippa, M. Eisner, D. Sulzer, S. Ito,
135. Wang, Y., and A. Casadevall. 1996. Susceptibility of melanized and non- R. Fariello, and M. Gallorini. 2002. The neuromelanin of human substantia
melanized Cryptococcus neoformans to the melanin-binding compounds
nigra and its interaction with metals. J. Neural Transm. 109:663–672.
trifluoperazine and chloroquine. Antimicrob. Agents Chemother. 40:541–
148. Zecca, L., D. Tampellini, M. Gerlach, P. Riederer, R. G. Fariello, and D. Sulzer. 2001. Substantia nigra neuromelanin: structure, synthesis, and mo-
136. Wasterstrom, S. A. 1984. Accumulation of drugs on inner ear melanin.
lecular behaviour. Mol. Pathol. 54:414–418.
Therapeutic and ototoxic mechanisms. Scand. Audiol. Suppl. 23:1–40.
137. Wasterstrom, S. A., and G. Bredberg. 1986. Ototoxicity of kanamycin in
149. Zhdanova, N. N., A. I. Gavriushina, and A. I. Vasilevskaia. 1973. Effect
albino and pigmented guinea pigs. II. A scanning electron microscopic
of gamma and UV irradiation on the survival of Cladosporium sp. and
study. Am. J. Otol. 7:19–24. Oidiodendron cerealis. Mikrobiol. Zh. 35:449–452.
138. Wheeler, M. H., and A. A. Bell. 1988. Melanins and their importance in
150. Zhdanova, N. N., and V. D. Pokhodenko. 1974. The protective properties of
pathogenic fungi. Curr. Top. Med. Mycol. 2:338–387.
fungal melanin pigment in some soil Dematiaceae. Radiat. Res. 59:221.
139. White, L. P. 1958. Melanin: a naturally occurring cation exchange material.
151. Zinn, K. M., and D. Z. Greenseid. 1975. Toxicology of the retinal pigment
Nature 182:1427–1428.
epithelium. Int. Ophthalmol. Clin. 15:147–158.
140. Wilkins, D. G., A. Mizuno, C. R. Borges, M. H. Slawson, and D. E. Rollins.
152. Zuger, A., E. Louie, R. S. Holzman, M. S. Simberkoff, and J. J. Rahal. 1986.
2003. Ofloxacin as a reference marker in hair of various colors. J. Anal.
Cryptococcal disease in patients with the acquired immunodeficiency syn-
Toxicol. 27:149–155.
drome: diagnostic features and outcome of treatment. Ann. Intern. Med.
141. Wrzesniok, D., E. Buszman, E. Karna, P. Nawrat, and J. Palka. 2002. 104:234–240.
Melanin potentiates gentamicin-induced inhibition of collagen biosynthesis
153. Zunino, H., and J. Martin. 1977. Metal-binding organic molecules in soil. 1.
in human skin fibroblasts. Eur. J. Pharmacol. 446:7–13.
Hypothesis interpreting the role of soil organic matter in the translocation
142. Wrzesniok, D., E. Buszman, E. Karna, and J. Palka. 2005. Melanin poten-
of metals ions from rocks to the biological system. Soil Sci. 123:65–76.
The Longwood Herbal Task Force The Center for Holistic Pediatric Education and Research Devil’s Claw ( Harpagophytum procumbens ) Principal Proposed Use: Anti-inflammatory for degenerative or rheumatic joint disease and Other Proposed Uses: Analgesic for other pains (headache, menstrual pain), antipyretic, antidiabetic, appetite stimulant and bitter tonic, liver and gall bladder
Dexamethasone for the Treatment of Sore Throat in Children With Suspected Infectious Mononucleosis A Randomized, Double-blind, Placebo-Controlled, Clinical Trial Michel Roy, MD, FRCPC; Benoit Bailey, MD, MSc, FRCPC; Devendra K. Amre, MBBS, PhD;Jean-Bernard Girodias, MD; Jean-Franc¸ois Bussie`res, BPharm, MSc, MBA, FCSHP; Pierre Gaudreault, MD, FRCPC Objective: To evaluate the efficacy
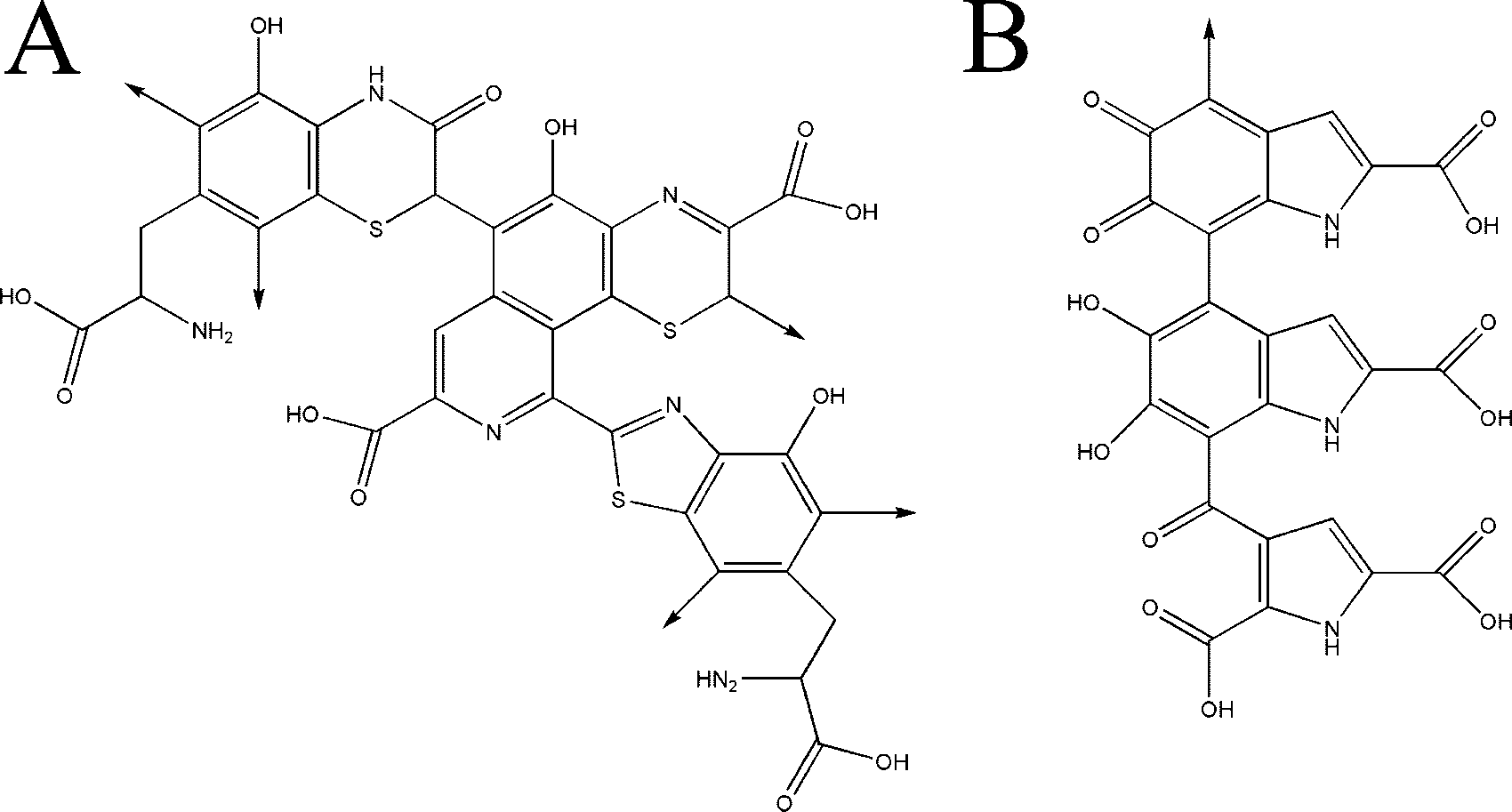 FIG. 1. Chemical structures of pheomelanin (A) and eumelanin (B) oligomers.
FIG. 1. Chemical structures of pheomelanin (A) and eumelanin (B) oligomers.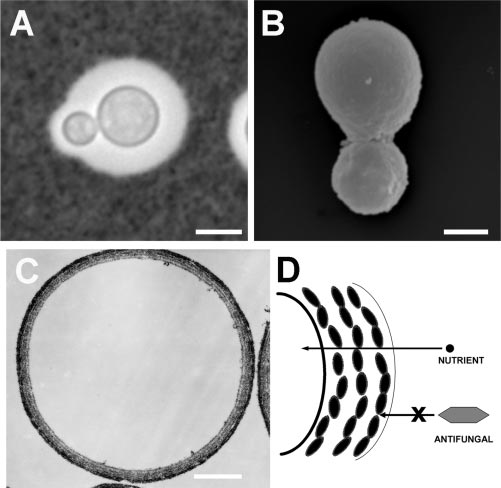 revealed highly specific melanin-binding sites for iodobenz-amides (11), which can be exploited to diagnose and stagemelanoma by using radiolabeled drug.
revealed highly specific melanin-binding sites for iodobenz-amides (11), which can be exploited to diagnose and stagemelanoma by using radiolabeled drug.