Kamagra enthält Sildenafilcitrat als pharmakologisch aktiven Bestandteil. Dieser hemmt selektiv die Phosphodiesterase-5 und erhöht dadurch die Konzentration von cGMP im Corpus cavernosum. Der Effekt ist zeitlich begrenzt, da die Halbwertszeit von Sildenafil etwa vier Stunden beträgt. In der galenischen Form als Mundgel erfolgt die Resorption besonders rasch, was zu einem schnelleren Wirkeintritt führt. Der Abbau erfolgt überwiegend hepatisch über CYP3A4, wobei ein aktiver Metabolit entsteht, der zur Gesamtwirkung beiträgt. Typische Nebenwirkungen ergeben sich aus der Vasodilatation, darunter leichte Kopfschmerzen und nasale Kongestion. In klinischen Beschreibungen wird kamagra oral jelly im Zusammenhang mit der schnelleren Absorption erwähnt.
Sansan.phy.ncu.edu.tw
Annu. Rev. Genet. 1997. 31:91–111Copyright c 1997 by Annual Reviews Inc. All rights reserved
GENE AMPLIFICATIONAND GENOMIC PLASTICITYIN PROKARYOTES
David Romero and Rafael PalaciosDepartment of Molecular Genetics, Nitrogen Fixation Research Center, NationalUniversity of Mexico, Apartado Postal 565-A, Cuernavaca, Morelos, Mexico;e-mail: dromero@cifn.unam.mx
genome structure, rearrangements, repeated DNA sequences, amplicons,adaptation
Gene amplification is a common feature of the genome of prokaryotic organisms. In this review, we analyze different instances of gene amplification in a varietyof prokaryotes, including their mechanisms of generation and biological role. Growing evidence supports the concept that gene amplification be considered notas a mutation but rather as a dynamic genomic state related to the adaptation ofbacterial populations to changing environmental conditions or biological interac-tions. In this context, the potentially amplifiable DNA regions impose a defineddynamic structure on the genome. If such structure has indeed been selectedduring evolution, it is a particularly challenging hypothesis.
INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 92GENOMIC STRUCTURE AND GENE AMPLIFICATION . . . . . . . . . . . . . . . . . . . . . . . . . . 92
The Canonical Model . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 92Extensions of the Canonical Model . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 95Amplifications in Streptomyces: A Different Paradigm . . . . . . . . . . . . . . . . . . . . . . . . . . . . 96Genome Structure and Dynamics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 97
MECHANISMS FOR GENE AMPLIFICATION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 100BIOLOGICAL ROLE OF GENE AMPLIFICATION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 103CONCLUSIONS AND PERSPECTIVES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 105
Gene amplification is a genetic phenomenon that is probably ubiquitous inthe genomes of prokaryotic organisms. The foundations for current inter-pretation of gene amplification were laid mainly through the elegant geneticwork of Roth, Hill, and their associates in Escherichia coli and in Salmonellatyphimurium (reviewed in 2, 37, 75, 93), as well as through studies by Rowndand his associates on R-factor “transitioning” in Proteus mirabilis (35, 73, 74). These studies allowed the establishment of what we call the canonical modelfor prokaryotic gene amplification. The quest for unity out of the diversityinherent in biological systems led to analysis of this phenomenon in otherprokaryotes. Initially recognized by purely genetic means, the application ofmolecular biological techniques to study amplification events in prokaryoteshas provided deeper insights. This work has expanded the basic tenets and hasextended the canonical model. However, because the characteristics of geneamplification in other organisms, with members of the genus Streptomyces asa prime example, do not fit so neatly into the canonical scheme, a differentparadigm emerges. Studies to define the biological role of gene amplificationhave provided a wealth of examples to illustrate the phenotypic richness thatcan be accrued through this phenomenon. The Canonical ModelThe pioneering studies on gene amplification in E. coli, S. typhimurium, andP. mirabilis were followed by a rapid expansion of knowledge about natural geneamplification in a wide range of prokaryotes. Most of the cases analyzed ad-here closely to the basic characteristics of the canonical model. Such examplesinclude Bacillus subtilis (34, 43, 67, 102, 108, 111, 113), Deinococcus radiodu-rans (62), Haemophilus influenzae (14, 45, 51, 100, 101), Klebsiella aerogenes(69), Pseudomonas aeruginosa (17), Rhizobium etli (27, 89, 90), Streptococcusfaecalis (109, 110), and Vibrio cholerae (31, 65). The central part of Figure 1(boxed) represents the basic features of the canonical model, as described below.
Most of the early work emphasized the astonishing feature that tandem du-
plications may affect almost any locus on the bacterial chromosome. However,the frequency of duplication for specific loci varies widely, from 10−2 to 10−5. Scanning of the whole S. typhimurium chromosome for the occurrence of dupli-cations, employing a single technique, revealed zones that behave as hotspotsin regard to tandem duplication, with frequencies as high as 3% (3). Hotspotshave been described in every bacterial system analyzed, whether chromosomalor plasmidic. A common feature in every characterized hotspot is the presence
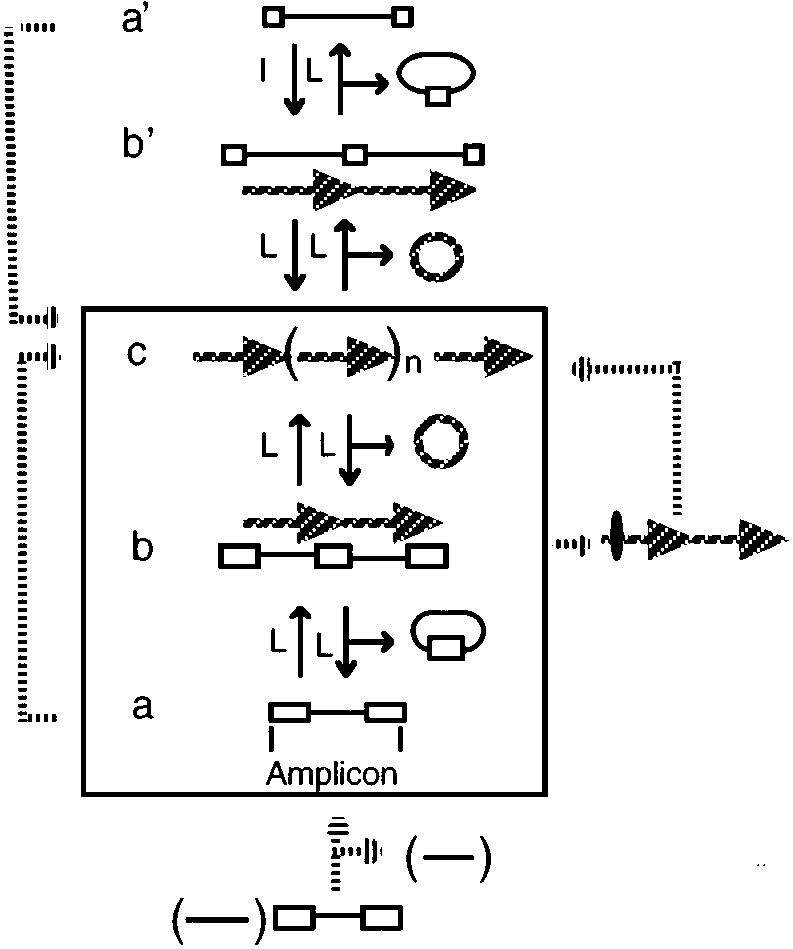
General models for gene amplification in prokaryotes. The central part (boxed ) illus-
trates the canonical model; alternate routes are shown outside the box, connected either by solid orbroken arrows. Most of the pathways shown here operate sequentially, leading to small increasesin copy number. However, routes shown in the left part might operate by bursts of amplificationleading to copy number increases of 100 or more. Lower-case letters indicate successive steps ineach case. Amplicon structures, a and a’; tandem duplications, b and b’; tandem amplification, c. Throughout the figure, long repeated sequences are shown as empty rectangles and short repeatsby empty squares. Long cross-hatched arrows represent the extent of tandem repeats formed; ovalon the cross-hatched arrow (right) represents a deletion. (L), legitimate, and (I) illegitimate recom-bination events. The bottom part shows an amplicon that is “activated” by deleting a large region(shown in parentheses) in the vicinity of the amplicon. The resulting amplicon is then amplifiedin a canonical way (central part) or by a sudden burst of amplification (left part).
of long repeated sequences (i.e. larger than 1 kb) in direct orientation, flankingthe region that undergoes tandem duplication. Structures composed of a DNAsegment bracketed by two repeated DNA sequences present in direct orienta-tion were recently dubbed “amplicons” (27, 90), to reflect the fact that they areprone not only to tandem duplication but to amplification as well (see below). The central part of Figure 1 (boxed) schematizes an amplicon structure (a), aswell as the structures generated during an amplification event. Homologousrecombination between the repeated sequences may lead either to a tandemduplication (Figure 1b) or to a deletion of the amplicon sequence (not shown).
These structures are thought to be formed by mechanisms entailing unequal ex-changes or through circle-excision and reinsertion (see Mechanisms for GeneAmplification). Tandem duplications generally do not lead to loss of gene func-tion (2, 93). Recombination between any pair of long repeated sequences, shar-ing an adequate level of identity, may possibly foster the formation of tandemduplications. Given the paucity of these sequences in enterobacterial genomes,chromosomal amplicons tend to be large—in some cases encompassing half ofthe chromosome of S. typhimurium (3). Size distribution may become skewedto smaller sizes in some prokaryotes that have a large abundance of repeatedsequences, such as Rhizobium (25) or Halobacterium (94).
The specific nature of the repeated sequences is unimportant in generating a
hotspot. Among the repeated sequences that may participate in generating tan-dem duplications, there are common inhabitants of many genomes, such as ri-bosomal (rrn) operons (3, 37, 38) and insertion sequences (IS) of different kinds(12, 32, 50, 74, 105, 109).
Examples of species-specific repeated sequences
that participate in amplification are the recombination hotspot sequences (rhs:Reference 58), found only in E. coli (114), or the repeated nitrogenase operonsin Rhizobium etli (89).
The main factors influencing ability to duplicate at high frequency are the
length and sequence conservation between the recombining repeats. Recom-bination between repeated sequences larger than 300 bp in E. coli is stronglydependent on recA, especially if those repeats are separated by a proportionallylong DNA segment (5). In this regard, null mutations in recA provoke a strongdecrease in the ability to generate tandem duplications (2, 31, 58, 75, 90, 98,110), whereas mutants harboring a constitutively activated recA allele showan enhanced duplication frequency (20). Small divergences in sequence (10%)between DNA repeats may reduce recombination frequency as much as 40-fold(97). Consequently, mutations in the major editor of genetic recombination inbacteria, the mismatch repair system (86), lead to an increase in tandem dupli-cation mediated by sequence-diverged repeats, such as the rhs elements (76). These constraints set an upper limit to the frequency of duplication of a givenamplicon.
The role of these constraints is relaxed after a tandem duplication is formed.
The large size and perfect homology between the halves of a tandem duplicationincrease the recombination frequency dramatically. Homologous recombina-tion between the tandemly duplicated sequences may lead either to furtheramplification (Figure 1c) or to deletion, resulting in the reversion to the origi-nal genomic structure, at frequencies as high as 150-fold over the frequency ofthe initial tandem duplication (2, 17, 31, 37, 74, 89, 90, 93). Successive recom-bination between these members leads to contractions and expansions in lengthof the tandem array. Thus, formation of amplifications under the canonical
model is a sequential process, where the rate-limiting step is the formation ofthe initial tandem duplication.
Interestingly, the dynamic nature of DNA amplification continuously gen-
erates closed circular structures that consist of monomers or multimers of theamplified unit (Figure 1). These structures will be lost owing to the lack of anorigin of replication (27, 38, 58, 100, 109). However, the use of some geneticmanipulations may allow such structures to be recovered (27, 58, 82). Thisprocess facilitates the molecular characterization of the amplicon sequences. Extensions of the Canonical ModelThe canonical model has been the paradigm for thinking about prokaryoticgene amplification. Its robustness is strengthened by the number of cases, indifferent prokaryotic species, that can be accurately explained by this model. Moreover, this model can be extended to explain and predict new instances. An interesting extension relates to the apparent stability of some tandem du-plications. Theoretical modeling indicates that, in the absence of selection forthe duplicated or amplified state, organisms harboring a single copy should be-come predominant in the population (47). Thus, if a stable tandem duplicationis found in a bacterial population, explanations invoking a positive selectivevalue for the duplication or some sort of recombinational avoidance must besought.
A fascinating example came from the analysis of H. influenzae genes in-
volved in capsule production. These genes are found naturally as part of atandem duplication in most of the isolates (39, 50, 51). “Stability” of the tan-dem duplication was achieved by deleting one copy of a gene essential forcapsulation (50). Reversal of the tandem duplication is frequently accompa-nied by loss of capsule production (39), which thus explains the predominanceof the duplication. According to the canonical model, generation of the tan-dem duplication is the limiting step for further amplification. Thus, this regionshould be “activated” for amplification. This prediction has been confirmed(14, 51). Such a situation is schematized in the right part of Figure 1.
A further extension to the canonical model is that duplications may also
be formed, albeit at much lower frequencies, by recombination between shortrepeated sequences (a’ in the upper part of Figure 1), for example, some tan-dem duplications in S. typhimurium, which can be formed through recombi-nation between the 35-bp long REP elements (98). Once the first duplicationis formed (Figure 1b), the remaining events that lead to amplification proceedthrough homologous recombination between the repeats. More extreme exam-ples, generated by illegitimate recombination (23) between shorter sequencerepeats (7–15 bp), have been detected spanning the hisG (98), the ampC (22),or the lacZ (104, 107) genes in E. coli.
More radical departures from the canonical model are suggested by the ex-
tremely high copy number that some amplifications may reach, at frequencies ashigh as 10−6 (12, 22, 44, 104, 107). These amplifications usually comprise smallamplicons, and they may establish at copy numbers as high as 100 (Figure 1,left). Because it is hard to reconcile such high-level amplification with thesequential steps and mechanisms currently invoked under the canonical model,other mechanisms (78), involving the use of rolling circle intermediates, havebeen considered.
Note that a particular locus in the genome may form part of different ampli-
cons. Different amplicons will show up depending on the nature of the test usedto detect or select the amplified variants. In general, amplicons with long andhighly similar direct repeats as borders will amplify at the highest frequency. These are usually large amplicons that give rise to a low number of tandemrepeats. When a large number of copies are demanded for a particular selectionprocedure, smaller amplicons will be detected. In some of these cases, the ini-tial duplication will occur at low frequency, mediated by a recA independent,illegitimate recombination. Amplifications in Streptomyces: A Different ParadigmExtreme deviations from the canonical model were found in Streptomycetes. Initial studies were motivated by the extreme instability, at frequencies usu-ally exceeding 10−2, exhibited by several important traits in these organisms. Among the unstable characters were aerial mycelium production, pigmenta-tion, sporulation, and/or antibiotic production. S. ambofaciens (55, 56) and S. lividans (6), instability is a biphasic phenomenon,where high instability for a certain trait is followed at frequencies as high as87% by instability for another trait. Frequencies of phenotypic instability canbe dramatically augmented by DNA-damaging treatments or by physiologicalstresses (6).
Genomic analysis of strains obtained from the first phase of instability re-
vealed massive chromosomal deletions, ranging from 250 to over 1500 kb insize (6, 54, 56), with no evidence for amplifications. In strains obtained fromthe second step of instability, these deletions may extend further, and they areusually accompanied by tandem amplification of specific chromosomal sectors(6, 54, 56), called AUDs (for Amplifiable Unit of DNA). Thus, amplificationevents in Streptomyces are part of a cascade of rearrangements. Available ev-idence indicates that amplification of the AUDs is either allowed or triggeredby large deletions (Figure 1, bottom), usually formed on only one side of theAUD (6, 54). After, or concomitant with, the deletion event, there is a burst ofamplification affecting the AUD (Figure 1, left). Copy number of the amplifiedregions is extremely high, reaching as much as 500 copies (6, 54). Depending
of the size of the amplified region, this copy number may represent as muchas 35% of the total DNA (6, 54). Amplifications in Streptomyces stand as themost dramatic example of gene amplification in any prokaryotic organism.
Two types of AUDs are recognized in Streptomyces. Type I AUDs usually
carry small (<25 bp long) repeated sequences in direct orientation (6, 36, 54,85); type II AUDs, are flanked by large (0.7–2.2 kb) direct repeats (6, 24, 87). These units bear a superficial resemblance to amplicons that duplicate by ille-gitimate or legitimate recombination, respectively.
Interestingly, some isolates of S. lividans harbor tandemly duplicated copies
of a type II AUD, whereas others possess a single copy (21). Isolates carrying asingle copy of the AUD amplify at much lower frequencies than do isolates har-boring the tandemly duplicated AUD (21). This might be explained assuming,as in the canonical model, that formation of a duplication is the rate-limitingstep for amplification. However, derivatives carrying amplifications of an AUDdeamplify very slowly (42). Circular structures, consisting of multimers of theAUD, have been observed in some species, such as S. ambofaciens (54). De-spite the superficial similarities with some structures postulated by the canonicalmodel, the idiosyncratic behavior of amplifications in Streptomyces imposes adifferent paradigm. Specific mechanisms for abrupt gene amplification of theAUDs and the connection with large deletions are considered in a later section.
An unexpected addition to an already alluring model was the recent finding
that the chromosomes of many Streptomyces species are linear (57, 59, 87). In both S. ambofaciens (54) and S. lividans (87), AUDs are located near theends of the chromosome, with deletions frequently removing one end (85). Interestingly, the genome switches from a linear to a circular conformation instrains harboring amplifications (87). These data give clear testimony of theenthralling possibilities for genomic plasticity in prokaryotes. Genome Structure and DynamicsThe potential for high-frequency DNA amplification in a genome depends to agreat extent on the amount, location, and relative orientation of long, repeatedsequences. These factors, coupled to the genomic architecture (chromosomalvs plasmidic, linear vs circular, reviewed in Reference 9) contribute to shapingthe specific “amplicon structure” of a particular genome.
The interplay between these factors creates some similarities but also strik-
ing differences, even between closely related genomes, such as E. coli andS. typhimurium (Figure 2). The most conspicuous families of long, reiteratedDNA elements in S. typhimurium are the rrn operons and the Salmonella-specific insertion sequence IS200. No other IS’s have been reported for thisorganism. A correlation exists between the location of these elements and theiruse to generate long duplications, some of them overlapping (32: reviewed in
Genomic maps of different bacterial replicons, showing the location of repeated families
and corresponding amplicons. Boxes and arrowheads represent repeated families, coded by shad-ing; uppercase letters on the R. etli map represent different families. Direction of the arrowheadsrepresent orientation of the respective element. In the maps of E. coli W3110, S. typhimurium LT2,and R. etli CFN42, experimentally demonstrated amplicons are represented by solid horizontallines; vertical lines mark the boundary for each amplicon. In the S. lividans 66 map, boxes onthe main map represent the unamplified AUDs. Beneath the map different rearrangements areshown; amplified regions are shown as boxes, and regions that are unaffected or deleted are shownas solid and broken lines, respectively. Information for this figure was collected from References37, 38, 58, 105 (E. coli); 2, 32, 93 (S. typhimurium); 30, 89, 90 (R. etli); and 57, 85, 87 (S. lividans).
93). In fact, most of the genome is subject to frequent duplication events; onlythe region surrounding the replication terminus is recalcitrant to duplication(93).
Amplicons of similar location and extent between E. coli and S. typhimurium
(Figure 2) are restricted to those involving recombination between the rrn oper-ons (37). In E. coli, amplicons may be bracketed by specific rhs sequences (58). Perhaps the most striking difference relates to the number of IS’s. E. coli haseight different types of IS’s, for a total of around fifty elements (18). In Figure 2we show the location of the most highly reiterated family, IS5. Specific pairs ofIS5 participate in the generation of tandem duplications (105). If most of theseelements participate in the formation of tandem duplications, the distributionof amplicons should be different between E. coli and S. typhimurium.
Highly dynamic genomes have been suggested for organisms that show a
distinctively high degree of DNA reiteration, such as Rhizobium (8, 26, 25, 91),Streptomyces (40), and Halobacterium (94, 95).
In Rhizobium, the genome is constituted by the chromosome and by large
circular plasmids ranging in size from 100 to 1700 kb. Plasmids may compriseup to 40% of the genome (61). In many Rhizobium species a single plasmid (thesymbiotic plasmid) carries most of the nodulation and nitrogen-fixation genes. Rhizobium genomes show a high reiteration, estimated for R. etli at 700 repeatedelements belonging to 200 distinct families (25). These repeated families arepresent in the chromosome and in the different plasmids. Amplicons have beenfound in the plasmids and in the chromosome of Rhizobium etli (27, 89, 90, 91)and Rhizobium tropici (P Mavingui & R Palacios, unpublished information).
The distribution of repeated sequences as well as the amplicons detected in
the symbiotic plasmid of R. etli are shown in Figure 2. This plasmid contains11 reiterated families containing 2–3 elements each (30). One of these familiescorresponds to the structural genes (nifH) for nitrogenase (83). Four ampliconswere detected in this plasmid, giving rise to duplications and higher-order am-plifications at frequencies ranging from 10−3 to 10−5 (89, 90). Interestingly,the amplicon bordered by the nif structural genes amplifies at the highest fre-quency. The four amplicons overlap extensively and range from 90 to 135kb (Figure 2). Partitioning of this plasmid into amplifiable and nonamplifiablezones is evident, since all the amplicons detected are concentrated in a zone ofroughly one third of the plasmid, covering most of the symbiotic genes (90). Asimilar case of partitioning was seen for the R-factor NR1 in Proteus mirabilis;this plasmid can be separated into amplifiable (the R-det) and nonamplifiable(RTF) regions (35, 73, 74).
Regions subject to deletion and amplification events in Streptomyces are
located close to the ends of the linear chromosome (Figure 2). Most of thedeletion events remove one end, although some remove both ends. Likewise,
amplification events frequently act on one AUD (85, 87). The reasons for thisparticular arrangement are unknown. The dispensability of the unstable regionfor growth under laboratory conditions was initially rationalized by assumingthat the unstable region contains relatively few genes. However, the existenceof profound changes in patterns of protein synthesis in S. ambofaciens strainsharboring extensive deletions indicates that these regions are actively expressed(16). A second alternative is that chromosome ends might play a role, as yetundescribed, on the generation of amplifications and deletions. In that regard, itshould be interesting to study the generation of amplifications in other bacterialspecies that contain linear chromosomes, such as Borrelia burgdorferi (10) andAgrobacterium tumefaciens (1).
The genomes of some archaebacteria also contain a large number of reiter-
ated DNA sequences and undergo frequent rearrangements (11, 94, 95). Thegenome of Halobacterium salinarium harbors an estimated 500 repeated ele-ments, belonging to as many as 50 different families (94). Most of this repeatedDNA consist of IS elements (11). Two genomic compartments of differing GC-content (FI and FII) have been found in the genome of H. salinarium (79). TheFII fraction is constituted by a large plasmid and by several long stretches (called“islands” in H. salinarium) of relatively AT-rich DNA embedded into GC-richchromosomal sequences. The IS elements are concentrated on the FII islands(79). Many rearrangements observed in this species are concentrated in the is-lands or are due to the movement of the IS to the FI fraction (33, 80, 81). Islands,also called “oases,” have been found also in Haloferax volcanii (13). Althoughfew examples of gene amplification have been described in haloarcheal species(92), it is expected that localization of the islands will play a role in definingthe amplicon organization in these species.
Figure 3 shows the different mechanisms proposed to explain the formationof gene amplification in prokaryotes. Two of these, the unequal crossover(UC) and the circle-excision and reinsertion (CER) mechanisms, might explainamplification under the canonical model (2, 37, 75, 93). The rolling circle (RC)mechanism was proposed to explain instances of abrupt gene amplification, asseen in Streptomyces (112) or in some “induced” amplifications in B. subtilis(78).
The UC and the CER mechanisms involve normal rounds of DNA replica-
tion; the UC mechanism in the form of a replication bubble, whereas the CERmechanism requires the existence of two replicons in the cell. Alternatively,the CER mechanism may use a replication bubble as an intermediate, excising acircle from an arm of the bubble and reinserting it into the other arm. In the UC
Mechanisms for gene amplification in prokaryotes. Repeated sequences are represented
by boxes of different shading; boxes with mixed shading represent the join points. Recombinationevents are represented as broken lines (see text).
mechanism, a reciprocal intermolecular recombination generates two products(a deletion and a duplication); a second, intramolecular recombination is neededto resolve the resulting dimeric molecule (Figure 3). Alternative proposals thatrecombination might proceed by half-reciprocal events would generate a fullproduct (a deletion or a duplication) and a broken molecule (60, 96). Therate-limiting step is the initial recombination; the second recombination eventapparently occurs as the site-specific recombination event usually involved inchromosome (7) or plasmid partitioning (15).
In the CER mechanism, a reciprocal intramolecular recombination generates
a deletion product and an excised circle; this circle then recombines with asecond molecule to produce the duplicated product. The efficiency of circleintegration may be very high (60); this can be explained by the extensive ho-mology between the circle and the recipient molecule, or by incorporating the
possibility of half-reciprocal events. A half-reciprocal recombination wouldgenerate a closed deletion product and a broken circle. Based on genetic evi-dence, Mahan & Roth (60) proposed that these linear molecules can be loadedby the RecBC enzyme; this promotes circularization, generating a circle nowactivated for integration, owing to the presence of RecBC.
The operation of the CER mechanism is made plausible by the demonstration
of circular intermediates in different systems (27, 38, 58, 100, 109). Studies onthe resolution of a nontandem duplication during spore formation in B. subtilisfailed to provide evidence for the UC mechanism, suggesting instead the CERmechanism (102). However, studies of resolution of a tandem duplication in aplasmid system in E. coli show that all the products in wild-type cells carried adimeric plasmid, containing a deletion and further amplification of the originaltandem duplication (19), supporting the UC mechanism. More studies areneeded to ascertain if there are species- or substrate-specificities for operationof these mechanisms.
Operation of the UC or CER mechanisms would lead to gradual increases in
copy number, requiring several generations to reach high-level amplification. Thus, these models are inadequate to explain sudden increases in copy number,as in Streptomyces. The RC mechanism (112) accounts nicely for the idiosyn-crasies of the Streptomyces model. This model proposes the entrapment of areplication fork into a circular structure that stays attached to a chromosomalarm. This structure would be generated by intramolecular recombination be-tween repeats in direct orientation, while the replication fork is traversing theAUD (Figure 3). A fork captured in such way would behave like a rolling circle,leading to the generation of tandem amplifications to a very high copy number. Resolution of this structure would be achieved by recombination between theoriginal repeats in the sister arm, generating an amplification without a deletion(Figure 3). Another alternative is by recombination downstream of the siteof the original recombination, generating an amplification accompanied by adeletion. This proposal is flexible enough to accommodate the possibility oflarge deletions prior to amplification, as seen in species that present biphasicinstability. These would occur through recombination, without trapping of areplication fork in the AUD structure. It is unclear if the ends of these linearchromosomes play a direct role in illegitimate recombination.
Operation of the RC mechanism was tested in Bacillus subtilis (77, 78) by
chromosomal integration of an activable, rolling circle–type origin in the vicin-ity of an artificial amplicon structure. Short-term activation of replication fromthis origin provoked amplification to a very high copy number, revealing atthe same time unstable intermediates predicted by the RC mechanism (77, 78). However, prolonged activation of replication leads to deletion of the array,probably due to enhanced recombination provoked by the generation of single-stranded DNA (71). Enhanced recombination was only seen with a rolling
circle–type origin in B. subtilis (66); however, recent results obtained with asimilar system in Rhizobium etli indicate that enhanced recombination mayoccur with a theta-type origin as well (E Valencia & D Romero, unpublishedresults).
Activation of a replication origin lying nearby might be involved in some
of the instances of sudden, high-level amplification observed previously. Forinstance, high-level amplification of the argF gene in E. coli occurs at highfrequency only in Hfr strains that transfer early the argF marker (12, 44). Con-ceivably, replication primed from the origin of conjugal transfer would generatea rolling circle–type structure, similar to the one seen in Figure 3. High-levelamplification of the lacZ gene in E. coli (104, 107) may well depend on a con-jugal transfer origin, since amplification was observed in strains harboring lacZon an F-prime plasmid. Similarly, tandem amplifications in Bacillus subtiliswere initially reported as exceedingly unstable, employing an experimental ap-proach that included an “artificial” amplicon (111). Elevated instability waslater shown to be dependent on the remnant of a rolling circle–type origin andits activating protein, inadvertently introduced during the construction of theartificial amplicon (113).
The life of bacteria has been colorfully described as “an adaptation to a feastor famine existence” (46). Genetic regulatory mechanisms in bacteria are nor-mally sufficient to cope with these demands. However, adaptation to new,extreme conditions or episodic situations may impose severe demands on theability of regulatory systems. It has been argued that the biological role of geneamplification in prokaryotes might be related to adaptation to these conditions(2, 88, 91, 93). According to this hypothesis, overexpression of gene productsthrough gene amplification may confer the phenotypic advantages needed forsurvival. The amplified state would remain as long as the selective conditionexists; when the selective condition disappears, haploid segregants would dom-inate the bacterial population. Different examples of natural gene amplificationsupport this proposal.
Several instances refer to increased resistance to antibiotics. A well-known
case is the amplification of the resistance determinant of plasmid NR1 inProteus vulgaris (35, 73, 74). Amplification of the R-det region (“transition-ing”) was seen upon growth of P. mirabilis cells harboring NR1 in mediumcontaining an antibiotic to which resistance is conferred by the R-det. In-creased antibiotic resistance correlates well with the degree of amplification. Amplification was mediated by recombination between IS1 elements flankingthe R-det. Deamplification, accompanied by a decrease in antibiotic resistance,was seen upon culturing in antibiotic-free medium (35, 73, 74).
Similarly, amplification of plasmid genes confers increased resistance to
tetracyclin in Streptococcus faecalis (109, 110). Amplification of specific chro-mosomal regions is also responsible for enhanced resistance in Escherichia colito ampicillin (22) or sulfonamides (70), and for increased resistance to methi-cillin in Staphylococcus aureus (63). In every case, deamplification leads todiminished antibiotic resistance.
Gene amplification might be involved in adaptation to other environmental
stresses, such as resistance to heavy metals. Thiobacillus ferrooxidans plays animportant role in the biogeochemical cycles of iron and sulfur. A recent reportsuggests that experimental adaptation for resistance to toxic metals (As and Zn)might be gained through amplification of chromosomal regions (48).
Adaptations for growth under conditions of nutrient scarcity can also be
achieved through gene amplification. Chemostat selection experiments demon-strate that tandem chromosomal duplications allow improved growth ofSalmonella typhimurium under limiting concentrations of malate (99, 103),arabinose, melibiose, or sorbitol (99). Improved permeation of the limitingsubstrate is partially responsible for this effect (99). Similar experiments inE. coli show that tandem duplications permit faster growth under limiting lac-tose concentrations, apparently due to increased synthesis of β-galactosidase(41, 104).
Growth on several “exotic” carbon sources can also be stimulated by gene
amplification. In this regard, growth of Klebsiella aerogenes in xylitol, anunnatural carbohydrate, is dependent on tandem amplification of the ribitoldehydrogenase gene region. Ribitol dehydrogenase has a very low affinityfor xylitol; thus, enzyme overproduction allows growth on this carbon source(69, 88). Bacteria belonging to the genera Alcaligenes and Pseudomonas pos-sess surprising catabolic activities toward several pollutants, including alkanesand halogenated compounds, such as 3-chlorobenzoate (3-Cba). Prolongedculturing of Alcaligenes eutrophus (28, 29, 72) or Pseudomonas sp. B13 (84)cells on media containing 3-Cba selects for variants carrying tandem amplifica-tion of the plasmid-encoded genes for 3-Cba-degrading enzymes. Tandem geneamplification has also been implicated in the phenotypic reversal of a mutationblocking alkane utilization in Pseudomonas putida (64).
Perhaps the best examples of the role of gene amplification in adaptive pro-
cesses came from studies of pathogenic or symbiotic interactions. Many isolatesof Vibrio cholerae, including epidemic strains, harbor a tandem duplication ofthe cholera toxin gene region (ctx). Interestingly, it was recently shown that ctxis part of a filamentous phage that generates the tandem duplication of ctx uponlysogenic conversion of V. cholerae (106). Further amplification of the ctx re-gion has been observed during the course of a mouse infection, correlating withincreased virulence (31, 65). Natural derivatives carrying amplifications havebeen isolated from humans (4). In a related example, many invasive strains of
Haemophilus influenzae carry a “stable” tandem duplication of genes involvedin capsule formation (39, 50, 51). One third of the H. influenzae strains isolatedfrom blood or cerebrospinal fluid of children with meningitis carry furtheramplification of the capsulation region (14). Increased capsulation affordedby gene amplification is thought to be an important factor for invasiveness(14, 51, 68). Another related example is associated with transition to mucoidyin Pseudomonas aeruginosa. In the course of infections by P. aeruginosa, bac-terial populations are frequently overtaken by mucoid, alginate-overproducerderivatives. These derivatives are one of the main causes of persistent infections. Although the basis for this transition is multifactorial, it has been demonstratedthat a chromosomal tandem amplification is implicated in this transition (17).
The benefits of gene amplification might be extended to other organisms, as
seen in symbiotic interactions. For instance, in certain genetic backgrounds,tandem duplication of a region in the symbiotic plasmid of Rhizobium etli,the symbiont of the bean plant, promotes increased nitrogen fixation in thishost (D Romero, unpublished data). Similarly, amplification of a region inthe symbiotic plasmid of Rhizobium tropici leads to enhanced production ofnodulation factors, which are necessary for nodule formation (P Mavingui &R Palacios, unpublished results). A further example is seen during the mutual-istic interaction of aphids with bacterial endosymbionts of the genus Buchnera. The main function of the endosymbiont is the supply of amino acids, speciallytryptophan, for the aphid. Genes encoding anthranilate synthase (trpEG) inBuchnera aphidicola are located in a plasmid, which is arranged as tandemrepeats of four to ten copies of trpEG (52). Thus, amplification of trpEG wouldprovide the means to overproduce tryptophan for the aphid host. This plasmid-encoded arrangement is prevalent in Buchnera species from aphids with a highdemand for tryptophan. Alternative arrangements are seen in endosymbiontsfrom aphids with conceivably low demand for tryptophan. These include singlechromosomal copies of trpEG or a plasmidic arrangement with a single activecopy of trpEG and six pseudogenes in tandem (53).
Gene amplification events provide several opportunities for the bacterium. First,tandem amplification endows a subset of the bacterial population with the ca-pability for overexpression of defined genomic regions. This capability mightbe relevant for short-term adaptation. Second, tandem amplification is a tran-sient characteristic, which may return to baseline conditions without alteringthe structure of the genome. Third, gene amplification provides not only im-mediate quantitative changes, but also new substrates for evolution. Amplifiedgenes can receive different mutations, thus allowing the evolution of new func-tions, while leaving the original copies unscathed. These opportunities allow
the bacterial genome to become truly experimental. In the short term, if aparticular gene amplification makes the bacterium best fitted for a particularenvironmental niche, whether episodic or entirely new, it will be strongly se-lected for. If not, there are two possible outcomes: Either the amplified regionreverts to the original state or the amplified copies may diverge by mutation. These opportunities might be particularly relevant for prokaryotic organisms,whose mode of reproduction is largely clonal.
A central biological question is whether the amplicon structure of a genome
is subjected to selection during evolution. No direct evidence exists at presentto confirm or rule out such a hypothesis: Arguments can be made both in favoror against it. In many cases a defined amplicon clearly confers a selectableadvantage to the organism. On the other hand, the amplicon structure of severalgenomes is strongly influenced by the position of mobile reiterated elementsand is thus continuously changing. Presumably, some amplicons have beenpositively selected during evolution whereas others are present as a collateralresult of evolutionary forces centered outside the amplicon itself.
Genome sequencing projects (see Reference 49 for a review) will certainly
provide new insights into the amplicon structure of the genome and its evolution. Analysis of the DNA sequence of a complete genome allows the predictionof potential amplicons contained in it, as well as their coding characteristics. Hypotheses on their biological role may then be derived and analyzed with thetools of modern molecular biology. Comparative genomics will help to definethe evolutionary history of genomes in terms of their amplicon structure.
We thank Daniel Balleza, Susana Brom, Marcela Castillo, Adriana Corvera,Patrick Mavingui, and Edgar Valencia for useful discussions, and AmparoGuti´errez and Jos´e Esp´ıritu for help in preparing the manuscript. Our workon gene amplification was partially supported by grants L0013N and 4321Nfrom CONACYT (M´exico). Visit the Annual Reviews home page at http://www.annurev.org.
teria. Annu. Rev. Microbiol. 31:473–
S, Jumas-Bilak E, Karayan L, Ramuz M.
1993. Presence of one linear and one cir-
cular chromosome in the Agrobacteriumtumefaciens C58 genome. J. Bacteriol.Salmonella typhimurium arise by unequal
(rrn) cistrons. Proc. Natl. Acad. Sci. USA
16. Dary A, Kaiser P, Bourget N, Thompson
in Vibrio cholerae O139. Microbiology
region in Streptomyces ambofaciens are
5. Bi X, Liu LF. 1994. recA-independent
gene expression. Mol. Microbiol. 10:759–
and recA-dependent intramolecular plas-
logy requirement and distance effect. J.
mucoid phenotype in rec-2 Pseudomo-
6. Birch A, H¨ausler A, H¨utter R. 1990. nas aeruginosa exposed to kanamycin. J.
bility in Streptomyces spp. J. Bacteriol.
18. Deonier RC. 1996. Native insertion se-
quence elements: locations, distributions
and sequence relationships. See Ref. 68a,
related recombinases are required for site-
specific recombination at dif and cer in E.coli K-12. Cell 75:351–61
anisms of deletion formation in Es-
8. Brom S, Garc´ıa de los Santos A, Girard
cherichia coli plasmids. I. Deletion for-
ML, D´avila G, Palacios R, Romero D.
mation mediated by long direct repeats. Rhizobium leguminosarum bv. phaseoli
20. Dimpfl J, Echols H. 1989. Duplication
plasmids. J. Bacteriol. 173:1344–46
mutation as an SOS response in Es-
tion in prokaryotes. Curr. Opin. Genet.
20a. Drlica K, Riley M, eds. 1990. The Bacte-rial Chromosome. Washington, DC: Am.
of Borrelia burgdorferi. Mol. Microbiol.
11. Charlebois RL, WF Doolittle. 1989. Streptomyces lividans 66. J. Bacteriol.
ture in halobacteria. In Mobile DNA, ed.
gies causes tandem duplication. Nature
tor in E. coli K12 is integrated in cis
Vilette D, Petranovic M, et al. 1993.
by IS1 repeats the elements are activated
so that amplification and other regulatory
changes that affect the gene can occur.
24. Eichenseer C, Altenbuchner J. 1994. The
from Streptomyces lividans 66 has inser-
tion sequence-like repeats at its ends. J.
of Haloferax volcanii, one of the archaea.
25. Flores M, Gonz´alez V, Brom S, Mart´ınez
Proc. Natl. Acad. Sci. USA 89:1602–6
E, Pi˜nero D, et al. 1987. Reiterated DNA
14. Corn PG, Anders J, Takala AK, K¨ayhty
sequences in Rhizobium and Agrobac-terium spp. J. Bacteriol. 169:5782–
Haemophilus influenzae type b capsule
expression are frequently amplified. J. In-
26. Flores M, Gonz´alez V, Pardo MA, Leija
A, Mart´ınez E, et al. 1988. Genomic insta-
15. Cornet F, Mortier I, Patte J, Louarn JM.
bility in Rhizobium phaseoli. J. Bacteriol.
bination site, psi, which is able to resolve
the analogous chromosomal Escherichia
amplification in Rhizobium: identifica-
coli site, dif. J. Bacteriol. 176:3188–
Rhizobium leguminosarum biovar phase-
an 18-kilobase tandem duplication. Proc.oli. Proc. Natl. Acad. Sci. USA 90:4932–
40. Hopwood DA, Kieser T. 1990. The Strep-tomyces genome. See Ref. 20a, pp. 147–
mids pJP4 and pJP2. Can. J. Microbiol.
41. Horiuchi R, Horiuchi S, Novick A. 1963.
The genetic basis of hypersynthesis of β-
galactosidase. Genetics 48:157–69
ing degradation of 3–chlorobenzoic acid
1989. DNA amplification in Streptomyces
in plasmids pAC27 and pJP4. Proc. Natl.achromogenes subsp. rubradiris is ac-
D, Palacios R, D´avila G. 1991. Structural
and can be eliminated by two pathways. Rhizobium leguminosarum bv. phaseoli.
43. Ives CL, Bott KF. 1990. Characteriza-
31. Goldberg I, Mekalanos JJ. 1986. Effect
with associated tetracycline resistance in
of a recA mutation on cholera toxin gene
Bacillus subtilis. J. Bacteriol. 172:4936–
amplification and deletion events. J. Bac-
44. Jessop AP, Clugston C. 1985. Amplifica-
tion of the argF region in strain HfrP4X of
E. coli K-12. Mol. Gen. Genet. 201:347–
mation in Salmonella typhimurium. Ge-
tion of DNA at a prophage attachment site
in Haemophilus influenzae. J. Bacteriol.
46. Koch AL. 1971. The adaptive responses
of Escherichia coli to a feast or famine
Halobacterium salinarium. J. Bacteriol.
existence. Adv. Microbial Physiol. 6:147–
47. Koch AL. 1979. Selection and recombi-
plification of the amyE-tmrB region on
multiple genes. J. Mol. Evol. 14:273–85
cells of Bacillus subtilis. Mol. Gen. Genet.
resistant strains of Thiobacillus ferroox-idans have increased copy numbers of
sition of the R factor NR1 in Proteus
chromosomal resistance genes. Microbi-mirabilis: level of drug resistance of non-
transitioned and transitioned cells. J. Bac-
49. Koonin EV, Mushegian AR, Rudd KE.
rial genomes. Curr. Biol. 6:404–16
50. Kroll JS, Loynds BM, Moxon ER. 1991.
nomic amplification in Streptomyces
The Haemophilus influenzae capsulation
glaucescens: structure, location and junc-
gene cluster: a compound transposon.
tion sequence analysis. Mol. Gen. Genet.
51. Kroll JS, Moxon ER, Loynds BM. 1993.
37. Hill CW, Harvey S, Gray JA. 1990.
in Escherichia coli and Salmonella ty-Haemophilus influenzae type b. J. Infect.phimurium. See Ref. 20a, pp. 335–39
52. Lai CY, Baumann L, Baumann P. 1994.
Amplification of trpEG: adaptation of
Buchnera aphidicola to an endosymbiotic
bosomal RNA genes in Escherichia coli.
association with aphids. Proc. Natl. Acad.
39. Hoiseth SK, Moxon ER, Silver RP. 1986.
53. Lai CY, Baumann P, Moran N. 1996. The
Genes involved in Haemophilus influen-
endosymbiont (Buchnera sp.) of the aphid
zae type b capsule expression are part of
Diuraphis noxia contains plasmids con-
trpEG pseudogenes. Appl. Environ. Mi-
homologous recombination in the Bacil-lus subtilis chromosome. Mol. Microbiol.
sights into the genetic instability of Strep-tomyces. FEMS Microbiol. Lett. 123:225–
fication on the Bacillus subtilis chro-
55. Leblond P, Demuyter P, Moutier L, Laakel
mosome. J. Gen. Microbiol. 134:85–
instability related to DNA amplification in
Streptomyces ambofaciens. J. Bacteriol.
56. Leblond P, Demuyter P, Simonet JM, De-
68a. Neidhardt FC, Curtiss R III, Ingraham
hypervariability in Streptomyces ambo-
1996. Escherichia coli and Salmonella ty-faciens: towards an understanding of a
phimurium: Cellular and Molecular Bi-
mechanism of genome plasticity. Mol.ology. Washington, DC: Am. Soc. Micro-
57. Leblond P, Redenbach M, Cullum J. 1993.
69. Neuberger MS, Hartley BS. 1981. Struc-
Physical map of the Streptomyces lividans
the related strain Streptomyces coelicolor
nase in a mutant of Klebsiella aerogenes.
A3(2). J. Bacteriol. 175:3422–29
plification contributes to sulfonamide re-
for duplications within the Escherichia
sistance in Escherichia coli. Antimicrob.coli K-12 chromosome. J. Mol. Biol.
71. Noirot P, Petit MA, Ehrlich SD. 1987.
combination in Bacillus subtilis. J. Mol.Streptomyces lividans 66 is linear. Mol.
60. Mahan MJ, Roth JR. 1989. Role of recBC
plasmid from Alcaligenes eutrophus NH9
mediated by direct repeat elements. Appl.
combination. Genetics 121:433–43
61. Mart´ınez E, Romero D, Palacios R. 1990.
73. Perlman D, Stickgold R. 1977. Selective
The Rhizobium genome. Crit. Rev. Plant
NR1, in Proteus mirabilis: an example of
the induction of selective gene amplifica-
tion. Proc. Natl. Acad. Sci. USA 74:2518–
Deinococcus radiodurans. J. Bacteriol.
resistance determinant DNA. J. Bacteriol.
DNA in methicillin-resistant Staphylo-coccus aureus following growth in high
concentrations of methicillin. J. Gen. Mi-
croorganisms. Annu. Rev. Genet. 22:147–
64. McBeth DL, Shapiro JA. 1984. Reversal
76. Petit MA, Dimpfl J, Radman M, Echols H.
tation blocking alkane and alcohol utiliza-
plications in Escherichia coli by the mis-
tion in Pseudomonas putida. Mol. Gen.
match repair system. Genetics 129:327–
77. Petit MA, Joliff G, Mesas JM, Klier A,
amplification of toxin genes in Vibrio
secretion of a cellulase from Clostridium
66. Morel-Deville F, Ehrlich SD. 1996. thermocellum in Bacillus subtilis by in-
the symbiotic plasmid of Rhizobium etli
Deville F, Ehrlich SD. 1992. Induction of
91. Romero D, D´avila G, Palacios R. 1997.
DNA amplification in the Bacillus subtilis
The dynamic genome of Rhizobium. In
Bacterial Genomes: Physical Structure
79. Pfeifer F, Betlach M. 1985. Genome or-
and Analysis. ed. FJ de Bruijn, JR Lup-
ganization in Halobacterium halobium: a
the chromosome. Mol. Gen. Genet. 198:
80. Pfeifer F, Blaseio U. 1989. Insertion
halophilic archaebacterium. J. Bacteriol.Halobacterium volcanii to the dihydrofo-
late reductase inhibitors trimethoprim and
81. Pfeifer F, Blaseio U. 1990. Transposition
methotrexate. Mol. Gen. Genet. 208:518–
burst of the ISH27 insertion element fam-
ily in Halobacterium halobium. Nucleic
93. Roth JR, Benson N, Galitski T, Haack K,
82. Posfai G, Koob M, Hradecn´a Z, Hasan
mation and applications. See Ref. 68a, pp.
segments of the Escherichia coli genome.
94. Sapienza C, Doolittle WF. 1982. Unusual
physical organization of the Halobac-
83. Quinto C, de la Vega H, Flores M, Lee-
terium genome. Nature 295:384–89
95. Sapienza C, Rose MR, Doolittle WF.
family in Rhizobium phaseoli. Proc. Natl.
peat sequence elements. Nature 299:182–
ability of Pseudomonas sp. strain B13
and deamplification of DNA. J. Bacteriol.
order chromosomal sequences. Genetics
85. Rauland U, Glocker I, Redenbach M, Cul-
97. Shen P, Huang HV. 1986. Homologous re-
combination in Escherichia coli: depen-
deletions in Streptomyces lividans 66 and
dence on substrate length and homology.
the loss of one end of the linear chromo-
some. Mol. Gen. Genet. 246:37–44
86. Rayssiguier C, Thaler DS, Radman M.
tween Escherichia coli and Salmonella
recombination event at the joint-point. typhimurium is disrupted in mismatch-
repair mutants. Nature 342:396–401
99. Sonti RV, Roth JR. 1989. Role of gene du-
87. Redenbach M, Flett F, Piendl W, Glocker
plications in the adaptation of Salmonella
I, Rauland U, et al. 1993. The Strepto-typhimurium to growth on limiting carbon
myces lividans 66 chromosome contains
100. Spies T, Laufs R. 1983. Circularized
two amplifiable regions. Mol. Gen. Genet.
from Haemophilus influenzae plasmids. J.
88. Rigby PWJ, Burleigh BD, Hartley BS.
101. Spies T, Laufs R, Riess FC. 1983.
enzyme evolution. Nature 251:200–4
89. Romero D, Brom S, Mart´ınez-Salazar J,
Haemophilus influenzae plasmids. J. Bac-
Girard ML, Palacios R, D´avila G. 1991.
Amplification and deletion of a nod-nif
region in the symbiotic plasmid of Rhizo-bium phaseoli. J. Bacteriol. 173:2435–41
erated sequences in the Bacillus subtilis
90. Romero D, Mart´ınez-Salazar J, Girard
crossing over. Biochimie 74:713–21
plifiable tetracycline resistance plasmid
in Salmonella typhimurium. Genetics
pAMa1 in Streptococcus faecalis. J. Bac-
104. Tlsty TD, Albertini AM, Miller JH. 1984.
110. Yagi Y, Clewell DB. 1980. Amplification
Gene amplification in the lac region of E.
of the tetracycline resistance determinant
of plasmid pAMa1 in Streptococcus fae-
105. Umeda M, Ohtsubo E. 1990. Mapping of
calis: dependence on host recombination
insertion element IS5 in the Escherichia
machinery. J. Bacteriol. 143:1070–72
coli K-12 chromosome. J. Mol. Biol.
in Bacillus subtilis. J. Gen. Microbiol.
106. Waldor MK, Mekalanos JJ. 1996. Lyso-
112. Young M, Cullum J. 1987. A plausible
phage encoding cholera toxin. Science
DNA amplification in streptomycetes.
113. Young M, Ehrlich SD. 1989. Stability of
reiterated sequences in the Bacillus sub-
in Escherichia coli: DNA sequences at
tilis chromosome. J. Bacteriol. 171:2653–
the junctures of amplified gene fusions.
114. Zhao S, Sandt CH, Feulner G, Vlazny DA,
Gray JA, Hill CW. 1993. Rhs elements
mo somal-DNA amplification in Bacillus
of Escherichia coli K-12: complex com-
subtilis. J. Bacteriol. 163:445–53
109. Yagi Y, Clewell DB. 1977. Identifica-
that have different evolutionary histories.
CONGRESSO NACIONAL DE ENFERMAGEM DE REABILITAÇÃO O ENFERMEIRO ESPECIALISTA E A SEXUALIDADE RESPOSTAS! Célia Mota – Enfª Especialista em Reabilitação – CoimbraO conceito de sexualidade humana engloba a forma como se pensa, sente e actua como um ser sexuado, com necessidades e impulsos, expressões de virilidade ou de feminilidade, papeis associados aos géneros, inter
RAMAH DAY CAMP in NYACK CAMPER Winter Address Summer Address PHOTO New York, NY 10027 PART I. (A parent or legal guardian should complete this page of the form) ______________________________________________________________________________________________ Camper Last Name ______________________________________________________________________________________________ Add
 General models for gene amplification in prokaryotes. The central part (boxed ) illus-
trates the canonical model; alternate routes are shown outside the box, connected either by solid orbroken arrows. Most of the pathways shown here operate sequentially, leading to small increasesin copy number. However, routes shown in the left part might operate by bursts of amplificationleading to copy number increases of 100 or more. Lower-case letters indicate successive steps ineach case. Amplicon structures, a and a’; tandem duplications, b and b’; tandem amplification, c.
General models for gene amplification in prokaryotes. The central part (boxed ) illus-
trates the canonical model; alternate routes are shown outside the box, connected either by solid orbroken arrows. Most of the pathways shown here operate sequentially, leading to small increasesin copy number. However, routes shown in the left part might operate by bursts of amplificationleading to copy number increases of 100 or more. Lower-case letters indicate successive steps ineach case. Amplicon structures, a and a’; tandem duplications, b and b’; tandem amplification, c.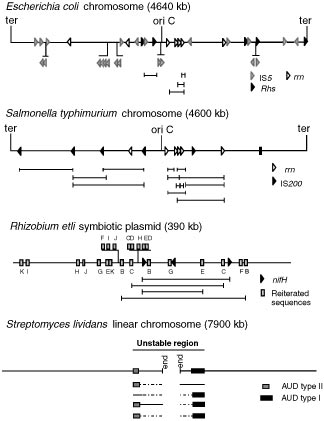 Genomic maps of different bacterial replicons, showing the location of repeated families
and corresponding amplicons. Boxes and arrowheads represent repeated families, coded by shad-ing; uppercase letters on the R. etli map represent different families. Direction of the arrowheadsrepresent orientation of the respective element. In the maps of E. coli W3110, S. typhimurium LT2,and R. etli CFN42, experimentally demonstrated amplicons are represented by solid horizontallines; vertical lines mark the boundary for each amplicon. In the S. lividans 66 map, boxes onthe main map represent the unamplified AUDs. Beneath the map different rearrangements areshown; amplified regions are shown as boxes, and regions that are unaffected or deleted are shownas solid and broken lines, respectively. Information for this figure was collected from References37, 38, 58, 105 (E. coli); 2, 32, 93 (S. typhimurium); 30, 89, 90 (R. etli); and 57, 85, 87 (S. lividans).
Genomic maps of different bacterial replicons, showing the location of repeated families
and corresponding amplicons. Boxes and arrowheads represent repeated families, coded by shad-ing; uppercase letters on the R. etli map represent different families. Direction of the arrowheadsrepresent orientation of the respective element. In the maps of E. coli W3110, S. typhimurium LT2,and R. etli CFN42, experimentally demonstrated amplicons are represented by solid horizontallines; vertical lines mark the boundary for each amplicon. In the S. lividans 66 map, boxes onthe main map represent the unamplified AUDs. Beneath the map different rearrangements areshown; amplified regions are shown as boxes, and regions that are unaffected or deleted are shownas solid and broken lines, respectively. Information for this figure was collected from References37, 38, 58, 105 (E. coli); 2, 32, 93 (S. typhimurium); 30, 89, 90 (R. etli); and 57, 85, 87 (S. lividans).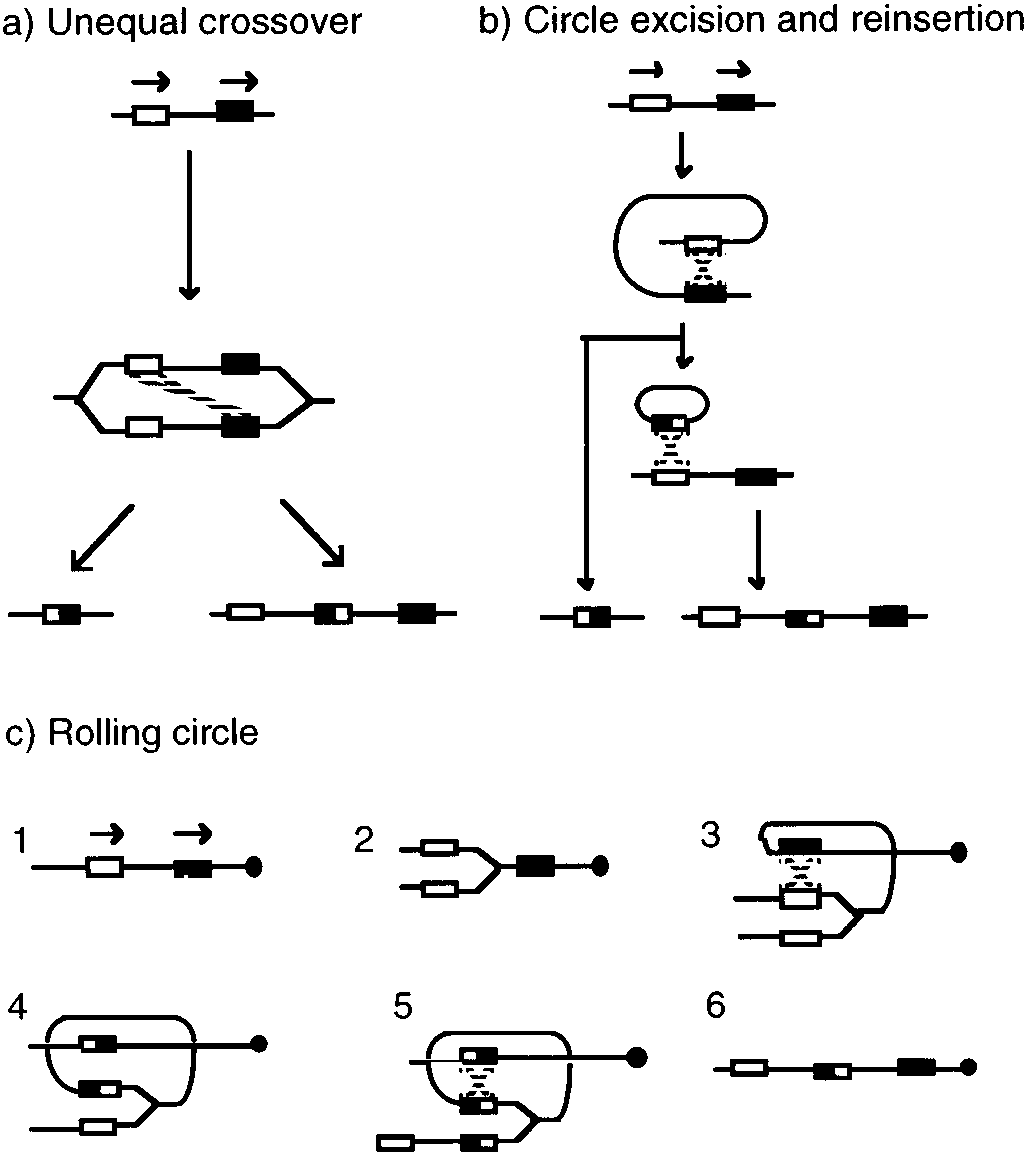 Mechanisms for gene amplification in prokaryotes. Repeated sequences are represented
by boxes of different shading; boxes with mixed shading represent the join points. Recombinationevents are represented as broken lines (see text).
Mechanisms for gene amplification in prokaryotes. Repeated sequences are represented
by boxes of different shading; boxes with mixed shading represent the join points. Recombinationevents are represented as broken lines (see text).