Kamagra enthält Sildenafilcitrat als pharmakologisch aktiven Bestandteil. Dieser hemmt selektiv die Phosphodiesterase-5 und erhöht dadurch die Konzentration von cGMP im Corpus cavernosum. Der Effekt ist zeitlich begrenzt, da die Halbwertszeit von Sildenafil etwa vier Stunden beträgt. In der galenischen Form als Mundgel erfolgt die Resorption besonders rasch, was zu einem schnelleren Wirkeintritt führt. Der Abbau erfolgt überwiegend hepatisch über CYP3A4, wobei ein aktiver Metabolit entsteht, der zur Gesamtwirkung beiträgt. Typische Nebenwirkungen ergeben sich aus der Vasodilatation, darunter leichte Kopfschmerzen und nasale Kongestion. In klinischen Beschreibungen wird kamagra oral jelly im Zusammenhang mit der schnelleren Absorption erwähnt.
Dkg268.fm
Journal of Antimicrobial Chemotherapy (2003) 52, 61–64 DOI: 10.1093/jac/dkg268 Advance Access publication 12 June 2003 Effect of chloramphenicol, erythromycin, moxifloxacin, penicillin and tetracycline concentration on the recovery of resistant mutants of Mycobacterium smegmatis and Staphylococcus aureus Tao Lu1, Xilin Zhao1, Xinying Li1, Glen Hansen2, Joseph Blondeau2 and Karl Drlica1* 1Public Health Research Institute, 225 Warren St., Newark, NJ 07103, USA; 2Departments of Clinical Microbiology, St. Paul’s Hospital (Grey Nuns’) and Saskatoon and District Health; Department of Pathology, Royal University Hospital and the Department of Microbiology and Immunology, University of Saskatchewan, Saskatoon, Saskatchewan, CanadaReceived 11 November 2002; returned 18 January 2003; revised 4 March 2003; accepted 2 April 2003The effect of antimicrobial concentration on colony-forming ability of resistant mutant subpopulations of Mycobacterium smegmatis and Staphylococcus aureus was measured for chloramphenicol, erythromy- cin, moxifloxacin, penicillin and tetracycline. The relationship between drug concentration and the recov- ery of mutant colonies was distinct for each bacterium–antimicrobial combination; however, in each case application of large numbers of cells to drug-containing agar plates revealed a progressive reduction in mutant recovery as antimicrobial concentration increased. The minimal concentration that allowed no mutant recovery from more than 1010 input cells was measured to estimate the minimum inhibitory concen- tration (MIC) of the least susceptible, single-step mutant subpopulation, a parameter also called the mutant prevention concentration (MPC). These data expand the number of antimicrobial–bacterial combinations for which a mutant selection window can be measured.
Keywords: erythromycin, moxifloxacin, penicillin, tetracycline, chloramphenicol
Introduction
always available for testing and since mutant susceptibility mightdiffer between pure cultures and small subpopulations, we have sug-
Surveillance studies with a variety of pathogens show that antimicro-
gested that MPC can be estimated as the concentration that allows
bial resistance can develop rapidly.1–8 If this trend is allowed to
recovery of no mutant when a large, susceptible population (1010 cells)
continue without the introduction of new classes of agent, manymicrobial diseases are likely to become refractory to antimicrobial
is applied to drug-containing agar plates.12 This microbiological
treatment. It may be possible to slow the development of de novo,
threshold can be readily measured in vitro for fluoroquinolones;12,13
step-wise resistance through more aggressive therapies that directly
however, it is not clear how the concept applies to agents of other
block the growth of the resistant mutant fraction of susceptible popu-
classes. For example, single-step mutations could lower suscep-
lations. In theory, antimicrobial therapy could be optimized such that
tibility so much that MPC could not be measured, as is the case with
failure due to the emergence of resistance during treatment occurs
only rarely.9 Empirical determination of such a dose is likely to
In the present work, we examined the recovery of resistant
require large numbers of patients to ensure that a particular regimen
mutants of Mycobacterium smegmatis and Staphylococcus aureus
does not contribute to the rising prevalence of resistance or an
from agar plates containing compounds representing five types of
increase in the frequency of other adverse events. As an alternative,
antimicrobial agent. In each case, the reduction in mutant recovery
we have proposed a conceptual strategy based on in vitro data.10,11 In
due to increasing drug concentration became progressively steeper
principle, antimicrobial concentrations that require a cell to attaintwo concurrent resistance mutations for growth will rarely allow
when large numbers of cells (109–1010) were applied to drug-contain-
selective enrichment of mutant subpopulations. That concentration is
ing agar. This is the result expected as antimicrobial concentration
the minimum inhibitory concentration (MIC) of the least susceptible,
approaches the MIC for the least susceptible single-step mutant,
single-step mutant, which is termed the mutant prevention concentra-
suggesting that MPC can be measured for antimicrobial agents of
tion (MPC). Since the least susceptible, single-step mutant is not
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
*Corresponding author. Tel: +1-973-854-3360; Fax: +1-973-854-3101; E-mail: drlica@phri.org
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
2003 The British Society for Antimicrobial Chemotherapy
T. Lu et al. Materials and methods Bacterial strains, growth conditions and antimicrobial agents
Wild-type M. smegmatis (strain mc2155), obtained from Dr S. Cole(Institute Pasteur, Paris, France), was grown at 37°C in 7H9 liquidmedium and on 7H10 agar, both supplemented with 10% albumen–dextrose complex (ADC).14 S. aureus strain RN450, obtained from DrB. Kreiswirth (Public Health Research Institute, Newark, NJ, USA), wasgrown at 37°C in CY broth (1% Casamino acids, 1% yeast extract, 0.1 MNaCl, 0.5% glucose and 0.05 M sodium glycerophosphate) and GL agar(0.3% Casamino acids, 0.3% yeast extract, 0.1 M NaCl, 0.2% sodiumlactate, 0.1% glycerol and 1.5% agar, pH 7.8).15 Bacteria were stored at–80°C in growth medium plus 15% glycerol. Chloramphenicol, erythro-mycin, penicillin and tetracycline were purchased from Sigma–AldrichCorp. (St Louis, MO, USA). Moxifloxacin was obtained from BayerCorp. (West Haven, CT, USA). Stock solutions (10 mg/mL) were pre-pared by dissolving penicillin in distilled water, moxifloxacin in 0.1 MNaOH, tetracycline in 50% ethanol, chloramphenicol in 95% ethanol,and erythromycin in 100% ethanol. Measurement of antimicrobial susceptibility
The minimum concentration that inhibited growth of 99% of the inputcells [MIC
] was measured by applying serial dilutions of stationary
phase cultures to agar plates containing various concentrations of anti-microbial agent. Colonies were counted after incubation (1–2 days forS. aureus; 3–4 days for M. smegmatis). Preliminary determinations usingtwo-fold dilutions of drug provided an approximate value of MIC
This measurement was followed by a second determination, plus a rep-licate, that utilized linear drug concentration increments (about 20% persequential increase). The fraction of colonies recovered was plottedagainst drug concentration to determine MIC(99) by interpolation.
MPC was defined as the concentration that blocked growth when at
least 1010 cells were applied to agar plates.11 To measure MPC, cells weregrown with vigorous shaking to reach a concentration of about 109 cfu/mL for M. smegmatis and 1010 for S. aureus. Cells were applied to drug-containing agar plates and, at the same time, the cell density of the culturewas determined retrospectively by applying serial dilutions to drug-freeagar. The maximal bacterial inoculum applied to each agar plate was300 µL of 1010 cfu/mL for S. aureus and 1 mL at 1 to 5 × 109 cfu/mL forM. smegmatis. Multiple plates at a given drug concentration were used sothat the total number of cells tested exceeded 1010. Agar plates were incu-bated at 37°C for various times depending on the species: 4 days withcolony numbers recorded at 1-day intervals for S. aureus and 10 dayswith colony numbers recorded at 2-day intervals for M. smegmatis. Colo-nies were confirmed to contain mutant cells by regrowth on agar contain-ing the selecting concentration of antimicrobial (control experiments inwhich mutant colonies were grown on drug-free agar before retesting ondrug-containing agar showed that the mutants were stable). M. smegmatis or S. aureus were plated on agar containing variousantimicrobial concentrations. After incubation, colonies werecounted and the proportion of the initial inoculum recovered as
Figure 1. Effect of antimicrobial concentration on recovery of resistant mutants.
colonies on the plates was calculated. As shown in Figure 1, raising
M. smegmatis strain mc2155 (open symbols) and S. aureus strain (RN450)
the antimicrobial concentration gave a sharp drop in colony recovery.
(filled symbols) were applied to agar plates containing the indicated concentra-
For some bacterium–antimicrobial combinations, a distinct shoulder
tions of (a) moxifloxacin, (b) erythromycin, (c) penicillin, (d) chloramphenicol
or plateau was observed in the recovery curves at high drug concen-
or (e) tetracycline. Triangles indicate concentrations at which no colony was
tration (moxifloxacin, erythromycin and tetracycline with M. smeg-
recovered when more than 1010 cells were applied to plates. Replicate experi-
matis; erythromycin with S. aureus). For others only inflection points
ments gave results similar to those shown. Effect of antimicrobials on recovery of resistant mutants Table 1. Mutant selection window for various antimicrobial agents with M. smegmatis and S. aureus aSpecific activity 1600 U/µg.
were detected (penicillin with M. smegmatis; moxifloxacin, tetra-
requires combination therapy even if the fraction of mutant cells is
cycline, penicillin and chloramphenicol with S. aureus). In the case
of chloramphenicol with M. smegmatis, no inflection point was
The second category of drug concentration dependence is illus-
trated by the combinations studied in the present work, by prior
Although the shape of the mutant recovery–drug concentration
studies with fluoroquinolones,12,18,19 and by treatment of Candida
curves varied among the bacterium–antimicrobial combinations, in
albicans and C. glabrata with miconazole (J.-Y. Wang et al., unpub-
each case the drop in mutant recovery became progressively steeper
lished observations). Increasing drug concentration causes colony
as antimicrobial concentration increased and high cell numbers were
recovery to drop sharply at the MIC, pass through an inflection point,
tested [the drop is probably steeper than shown, since the drug
and then drop sharply a second time. The second drop occurs at the
concentrations at which no colony was recovered (triangles) over-
MPC. Fluoroquinolone studies show that resistant mutants are selec-
estimate the true drug concentrations required to block colony
tively enriched at drug concentrations between MIC
growth]. Increasing steepness is the result expected as the MIC for the
MPC.16,18 Whether monotherapy is appropriate for situations in this
least susceptible mutant (MPC) is approached.11
category depends on how long relevant tissue drug concentrations
It has been argued that resistant mutants are selectively enriched
can be kept above MPC at each dosing interval. To address this issue,it is now necessary to measure MPC in vivo at the site of infection.
when antimicrobial concentrations fall between the minimal concen-
Confusion sometimes surrounds phenotypic or induced resist-
tration that inhibits the growth of 99% of the cells [MIC
ance. An example is the β-lactam resistance that arises from the
a range called the mutant selection window.10,13,16 [MIC
induction of β-lactamases. When mutations are not responsible for
mates the minimal concentration better than MIC because less select-
this type of resistance, it is outside the scope of the present discussion.
ive pressure is present; however, for many antimicrobial–pathogen
However, when β-lactamases are expressed from plasmid-borne
combinations, little absolute difference is likely to exist between
genes, they behave as category-one resistance and require com-
and MIC due to the steep dependence of colony recovery on
bination therapy even if the plasmid-containing cells are members of
rare subpopulations. Thus the dosing strategies that derive from con-
calculated and are listed in Table 1. When the size of the selection
sideration of the mutant selection window hypothesis may be broadly
window was expressed as the ratio of MPC to MIC
siderably among bacterial–antimicrobial combinations. For bothbacterial species, the selection window was widest for erythromycin. Acknowledgements Discussion
We thank Richard Burger and Marila Gennaro for critical commentson the manuscript prior to submission. The work was supported by
Although the relationship between antimicrobial concentration and
grants from the NIH (AI35257) and Bayer AG.
growth of bacterial mutants on drug-containing agar is characteristicof each antimicrobial–bacterium combination, two general catego-
References
ries can be identified. One, represented by rifampicin treatment ofseveral organisms, exhibits a sharp drop in colony recovery followed
1. Marcus, N., Peled, N. & Yagupsky, P. (1997). Rapid increase in
by a broad plateau as drug concentration increases.17 This result is
the prevalence of antimicrobial drug resistance among enterococcal
most easily explained by resistance mutations reducing susceptibility
blood isolates in southern Israel. European Journal of Clinical Micro-
so much that no achievable drug concentration can inhibit mutant
biology and Infectious Diseases 16, 913–5. 2. Glupczynski, Y. (1998). Antimicrobial resistance in Helicobacter
growth. A similar result is expected from a bacterial population
pylori: a global overview. Acta Gastroenterologica Belgica 61, 357–66.
having a subpopulation of plasmid-containing cells that exhibit high-
3. Gorwitz, R. K., Nakashima, A. K., Moran, J. S. & Knapp, J. S.
level resistance. In these situations no monotherapy regimen will
(1993). Sentinel surveillance for antimicrobial resistance in Neisseria
keep drug concentrations above MPC and thereby block mutant
gonorrhoeae—United States, 1988–1991. Morbidity and Mortality Weekly
growth. Restricting the development of this type of resistance
Report 42, 29–39. T. Lu et al. 4. Gallardo, F., Ruiz, J., Marco, F., Towner, K. J. & Vila, J. (1999). 12. Dong, Y., Zhao, X., Domagala, J. & Drlica, K. (1999). Effect of
Increase in incidence of resistance to ampicillin, chloramphenicol, and
fluoroquinolone concentration on selection of resistant mutants of Myco-
trimethoprim in clinical isolates of Salmonella serotype Typhimurium
bacterium bovis BCG and Staphylococcus aureus. Antimicrobial Agents
with investigation of molecular epidemiology and mechanisms of resist-
and Chemotherapy 43, 1756–8.
ance. Journal of Medical Microbiology 48, 367–74. 13. Zhao, X. & Drlica, K. (2002). Restricting the selection of antibiotic- 5. Johnson, A. P. (1998). Antibiotic resistance among clinically
resistant mutants: measurement and potential uses of the mutant selec-
important Gram-positive bacteria in the UK. Journal of Hospital Infection
tion window. Journal of Infectious Diseases 185, 561–5. 40, 17–26. 14. Jacobs, W. R., Kalpana, G. V., Cirillo, J. D., Pascopella, L., Snap- 6. Rudolph, K. M., Parkinson, A. J., Reasonover, A. L., Bulkow, L. R.,
per, S. B., Udani, R. A. et al. (1991). Genetic systems in mycobacteria.
Parks, D. J. & Butler, J. C. (2000). Serotype distribution and antimicrobial
Methods in Enzymology 204, 537–55.
resistance patterns of invasive isolates of Streptococcus pneumoniae:
15. Novick, R. P. & Brodsky, R. (1972). Studies on plasmid repli-
Alaska, 1991–1998. Journal of Infectious Diseases 182, 490–6.
cation. I. Plasmid incompatibility and establishment in Staphylococcus
7. Bennish, M., Salam, M. A., Hossain, M. A., Myaux, J., Khan, E.
aureus. Journal of Molecular Biology 68, 285–302.
H., Chakraborty, J. et al. (1992). Antimicrobial resistance of Shigella iso-
16. Firsov, A. A., Vostrov, S. N., Lubenko, I. Y., Drlica, K., Portnoy,
lates in Bangladesh, 1983–1990: increasing frequency of strains multi-
Y. A. & Zinner, S. H. (2003). In vitro pharmacodynamic evaluation of the
ply resistant to ampicillin, trimethoprim–sulfamethoxazole, and nalidixic
mutant selection window hypothesis using four fluoroquinolones against
acid. Clinical Infectious Diseases 14, 1055–60.
Staphylococcus aureus. Antimicrobial Agents and Chemotherapy 47, 8. Bal, C. (1999). Increasing antimicrobial resistance in STDs and
the need for surveillance: Neisseria gonorrhoeae as a model. FEMS
17. Dong, Y., Zhao, X., Kreiswirth, B. & Drlica, K. (2000). Mutant pre-
Immunology and Medical Microbiology 24, 447–53.
vention concentration as a measure of antibiotic potency: studies with
9. Thomas, J., Forrest, A., Bhavnani, S., Hyatt, J. M., Cheng, A.,
clinical isolates of Mycobacterium tuberculosis. Antimicrobial Agents
Ballow, C. H. et al. (1998). Pharmacodynamic evaluation of factors
and Chemotherapy 44, 2581–4.
associated with the development of bacterial resistance in acutely ill
18. Zhou, J.-F., Dong, Y., Zhao, X., Lee, S., Amin, A., Ramaswamy,
patients during therapy. Antimicrobial Agents and Chemotherapy 42,
S. et al. (2000). Selection of antibiotic resistance: allelic diversity among
fluoroquinolone-resistant mutations. Journal of Infectious Diseases 182, 10. Zhao, X. & Drlica, K. (2001). Restricting the selection of antibiotic-
resistant mutants: a general strategy derived from fluoroquinolone stud-
19. Blondeau, J., Zhao, X., Hansen, G. & Drlica, K. (2001). Mutant
ies. Clinical Infectious Diseases 33, Suppl. 3, S147–56.
prevention concentrations (MPC) of fluoroquinolones for clinical isolates
11. Drlica, K. (2003). The mutant selection window and antimicrobial
of Streptococcus pneumoniae. Antimicrobial Agents and Chemotherapy
resistance. Journal of Antimicrobial Chemotherapy 52, 11–7. 45, 433–8.
O TRABALHO E A SAÚDE NA CULTURA CONTEMPORÂNEAApresentarei, de início, uma oposição entre o Direito e a Psicanálise. Opensamento jurídico é permeado por uma pergunta: O que leva um homem atornar-se antissocial? As formalizações psicanalíticas, por sua vez, também sãoinstigadas por uma pergunta, que, entretanto, é o inverso da primeira: O que levaum homem a tornar-se social? A difere
STIPENDIARY STEWARDS REPORT Canberra Racing Club Incorporated THOROUGHBRED PARK Friday 13 May 2011 Weather: OvercastTrack: Good (3)Rail: TrueJ. D. Walshe (Chairman) C. P. Yeo, C. Polglase (Stewards), B. Delaney (Starter), P Selmes (Assistant Starter), R. Staggard (Betting), M. Kay (Swab), S. Farrar (Judge), R. Joyce (Veterinarian). ____________________________________________________
 Journal of Antimicrobial Chemotherapy (2003) 52, 61–64
Journal of Antimicrobial Chemotherapy (2003) 52, 61–64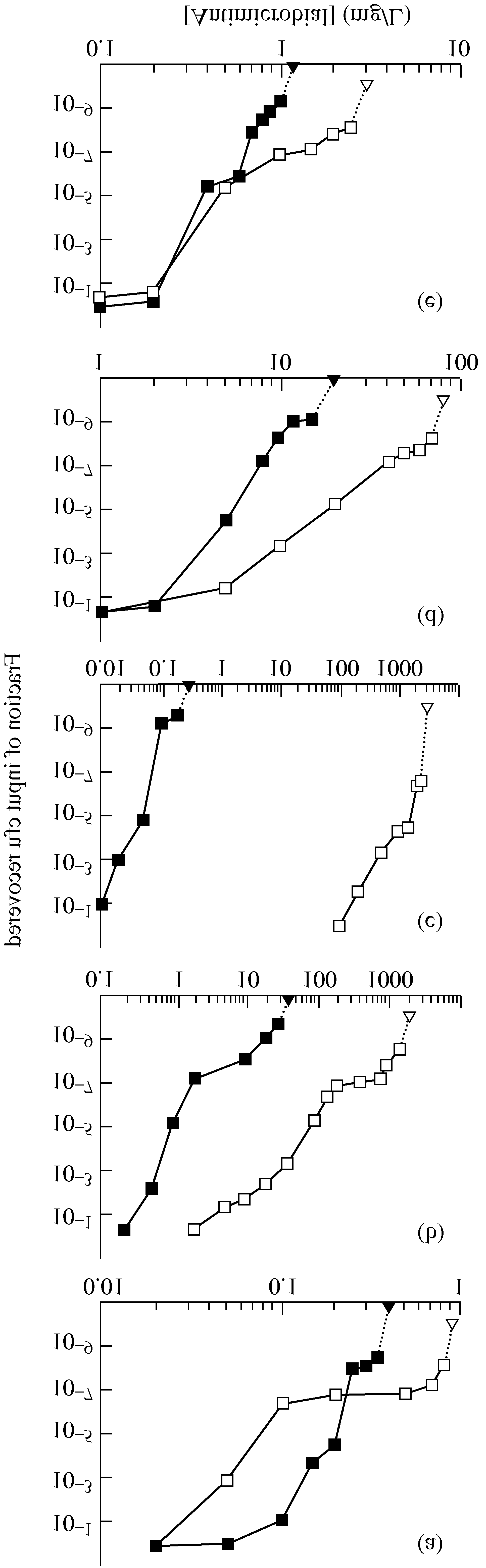 T. Lu et al.
T. Lu et al.