Kamagra enthält Sildenafilcitrat als pharmakologisch aktiven Bestandteil. Dieser hemmt selektiv die Phosphodiesterase-5 und erhöht dadurch die Konzentration von cGMP im Corpus cavernosum. Der Effekt ist zeitlich begrenzt, da die Halbwertszeit von Sildenafil etwa vier Stunden beträgt. In der galenischen Form als Mundgel erfolgt die Resorption besonders rasch, was zu einem schnelleren Wirkeintritt führt. Der Abbau erfolgt überwiegend hepatisch über CYP3A4, wobei ein aktiver Metabolit entsteht, der zur Gesamtwirkung beiträgt. Typische Nebenwirkungen ergeben sich aus der Vasodilatation, darunter leichte Kopfschmerzen und nasale Kongestion. In klinischen Beschreibungen wird kamagra oral jelly im Zusammenhang mit der schnelleren Absorption erwähnt.
No job name
Biochemistry 1999, 38, 3067-3072
Stabilization of Tubulin by Deuterium Oxide†
Gopal Chakrabarti,‡ Shane Kim,‡ Mohan L. Gupta, Jr.,‡ Janice S. Barton,§ and Richard H. Himes*,‡
Department of Molecular Biosciences, UniVersity of Kansas, Lawrence, Kansas 66045, and Department of Chemistry,Washburn UniVersity, Topeka, Kansas 66621ReceiVed October 15, 1998; ReVised Manuscript ReceiVed January 21, 1999
ABSTRACT: Tubulin is an unstable protein when stored in solution and loses its ability to form microtubulesrapidly. We have found that D2O stabilizes the protein against inactivation at both 4 and 37 °C. In H2O-based buffer, tubulin was completely inactivated after 40 h at 4 °C, but in buffer prepared in D2O, noactivity was lost after 54 h. Tubulin was completely inactivated at 37 °C in 8 h in H2O buffer, but only20% of the activity was lost in D2O buffer. Tubulin also lost its colchicine binding activity at a slowerrate in D2O. The deuterated solvent retarded an aggregation process that occurs during incubation at bothtemperatures. Inactivation in H2O buffer was partially reversed by transferring the protein to D2O buffer;however, aggregation was not reversed. The level of binding of BisANS, a probe of exposed hydrophobicsites in proteins, increases during the inactivation of tubulin. In D2O, the rate of this increase is slowedsomewhat. We propose that D2O has its stabilizing effect on a conformational step or steps that involvethe disruption of hydrophobic forces. The conformational change is followed by an aggregation processthat cannot be reversed by D2O. As reported previously [Ito, T., and Sato, H. (1984) Biochim. Biophys. Acta 800, 21-27], we found that D2O stimulates the formation of microtubules from tubulin. We alsoobserved that the products of assembly in D2O/8% DMSO consisted of a high percentage of ribbonstructures and incompletely folded microtubules. When these polymers were disassembled and reassembledin H2O/8% DMSO, the products were microtubules. We suggest that the combination of D2O and DMSO,both stimulators of tubulin assembly, leads to the rapid production of nuclei that lead to the formation ofribbon structures rather than microtubules.
Microtubules are cytoskeletal structures in eukaryotic cells
bovine brain by two cycles of polymerization and depolym-
that are involved in various types of motility processes,
erization (2). Phosphocellulose chromatography was used to
including chromosome positioning and separation during
remove the associated proteins (3), and the protein was stored
mitosis and meiosis, and vesicle transport. The protein
in PEM buffer1 as frozen pellets at -70 °C.
subunit of microtubules, tubulin, is an interesting protein that
For the stability and aggregation experiments described
binds nucleotides and a variety of antimitotic compounds,
below, it was necessary to obtain tubulin in D2O buffer. To
some of which are used as antitumor agents. Tubulin is
accomplish this, tubulin was thawed and centrifuged at
known to be an unstable protein, losing its microtubule-
40000g for 10 min to remove any aggregated material. The
forming activity over a period of hours when stored in
supernatant was made 8% in DMSO (4), 1 mM in DTT,
solution at low temperatures. The loss in activity is ac-
and 0.5 mM in GTP and was incubated at 37 °C for 15 min
companied by an irreversible aggregation process that
to form microtubules. The suspension was divided into two
involves structural changes resulting in changes in the
portions that were centrifuged in a Beckman TL-100 ultra-
fluorescence and circular dichroism spectra of the protein
centrifuge at 100000g for 4 min at 37 °C. The pellets were
(1). This instability of tubulin has hindered biochemical and
suspended in PEM buffer containing 1 mM DTT that had
biophysical studies that involve long time periods. In the
been prepared with either H2O or D2O. The D2O concentra-
course of conducting long-term NMR experiments with
tion in the PEM buffer prepared in D2O was 89%. After 30
tubulin, we found that D2O has a dramatic stabilizing effect
min on ice, the solutions were centrifuged at 40000g for 5
on the protein. This effect is examined in more detail in this
min. The protein concentrations of the supernatants were
report. In addition, we report the partial reversal of the
determined by the Bradford method (5), and the concentra-
tions were adjusted with buffer, if necessary, so that theywere equivalent. EXPERIMENTAL PROCEDURES Assembly Assays. Assembly reactions were carried out in
Purification of Tubulin. Microtubule protein (tubulin
100 µL volumes containing 1.5 mg/mL tubulin, PEM buffer
containing microtubule-associated proteins) was isolated from
prepared in H2O or D2O, 1 mM DTT, 8% DMSO, and 0.5
mM GTP. After 15 min at 37 °C, the samples were
This research was supported by NIH Grant CA 55141. G.C. was
a recipient of a Kansas Health Foundation Postdoctoral Cancer ScholarStipend.
1 Abbreviations: PEM buffer, 0.1 M 1,4-piperazinediethanesulfonate,
* To whom correspondence should be addressed. Telephone: (785)
1 mM ethylene glycol bis( -aminoethyl ether)-N,N,N′,N′-tetraacetic
864-3813. Fax: (785) 864-5294. E-mail: himes@kuhub.cc.ukans.edu.
acid, and 1 mM MgSO4; H2O buffer, PEM buffer prepared in H2O;
D2O buffer, PEM buffer prepared in D2O; BisANS, 5,5′-bis(8-anilino-
1-naphthalenesulfonate); DTT, dithiothreitol.
3068 Biochemistry, Vol. 38, No. 10, 1999
centrifuged at 100000g for 4 min at 37 °C . The pellet wassuspended in cold buffer and centrifuged at 40000g for 5min at 4 °C. After the supernatant was removed, 100 µL of0.1 M NaOH was added to dissolve any cold-insolublepellets. Protein concentrations of both supernatants and thecold-insoluble pellets were determined. In some cases, theassembly was monitored by the increase in turbidity at 350nm in reactions carried out in 400 µL volumes. Colchicine Binding Assay. The increase in colchicine
fluorescence that occurs when the drug binds to tubulin (6)was used to follow the loss in colchicine binding activity. Samples of tubulin were diluted with PEM buffer to aconcentration of 5 µM, and colchicine was added to aconcentration of 10 µM. The samples were incubated at 37
C for 1 h, and the fluorescence at 430 nm was read against
Tubulin at 1.5 mg/mL was assembled at 37 °C in the presence of
10 µM colchicine using an excitation wavelength of 350 nm.
0.5 mM GTP. (1) Assembled in H2O buffer with 8% DMSO. (2)
Measurement of Aggregation. The formation of aggregates
Assembled in D2O buffer without DMSO. (3) Assembled in D2Obuffer with 8% DMSO. (4) Tubulin was assembled in D
was monitored by size-exclusion HPLC using a 600 mm ×
containing 8% DMSO; microtubules were collected by centrifuga-
7.8 mm Phenomenex Biosep-Sec-2000 column. Tubulin
tion, suspended in H2O buffer, and repolymerized at 1.5 mg/mL in
samples were incubated at either 4 or 37 °C at a concentration
of 1 mg/mL. At different times, 250 µL aliquots wereremoved and applied to the column. In the case of the 37
development, the increase observed in buffer containing D2O
°C incubation, the samples were placed on ice for 15 min to
was consistently about twice that which developed in buffer
depolymerize any microtubules that may have formed and
containing H2O (Figure 1). This result suggested that a
centrifuged to remove cold-insoluble polymers before injec-
greater degree of assembly occurred in the D2O buffer.
tion. PEM buffer was used as the elution solution at a flow
However, when samples were centrifuged, there was no
rate of 1.0 mL/min. Detection was at 280 nm. Blue dextran,
significant difference in the supernatant or cold-soluble pellet
tryptophan, and a Bio-Rad protein gel filtration standard
protein concentrations. In both cases, about 75% of the
mixture consisting of protein aggregates, thyroglobin (670
protein had assembled into cold-soluble products. When
kDa), IgG (150 kDa), ovalbumin (44 kDa), myoglobin (17
DMSO was omitted from the assembly reaction solution
12 (1.35 kDa) were used as standards.
2O, the absorbance was reduced to the level seen
Areas under the aggregate and tubulin dimer peaks were
for the H2O/8% DMSO solution (Figure 1).
calculated using the NIH Image 1.60 program.
Since the degree of turbidity depends on the nature of the
Electron Microscopy. Negative staining was used to
structure present, the results suggest a difference in the
observe the structures of the polymerized products in the
structure of the polymer formed in D2O buffer containing
assembly reactions. Samples were diluted 10- or 20-fold into
DMSO. Examination of negatively stained samples showed
PEM buffer containing 0.25% glutaraldehyde, applied to 300-
that this was indeed the case. In the case of assembly in
mesh carbon-coated copper grids, stained with 2% uranyl
either H2O/8% DMSO or D2O without DMSO, the polymers
acetate, and viewed with a JEOL JEM-1200 EX11 electron
were essentially all microtubules (Figure 2A,B), but a high
percentage of structures formed in D2O/8% DMSO were
open ribbons and incompletely closed microtubules (Figure
2O concentration of a solution after dialysis
2O solution was determined by 1H NMR using a
Brucker Advance DRX 400 spectrometer operating at 400.13
Because there are differences between the acidity of D2O
MHz. The isotopic composition of the dialysate was deter-
and H2O, the chemical activities of D+ and H+, and pKa
values of weak acids in D2O and H2O, the pH meter reading
peak resulting from the addition of a known quantity of
of a solution in D2O must be corrected with the relationship
pD ) pH meter reading + 0.4 (7). The pH meter readingsof the buffer solutions in the experiments described in Figures
Materials. D2O (99.9%) was purchased from Cambridge
1 and 2 were 6.9. To ensure that the difference in pH and
Isotope Laboratories, Inc. BisANS was from Molecular
pD did not contribute to the results, we also conducted
Probes, Inc. GTP was obtained from Boehringer Mannheim.
assembly reactions in buffers with a pH meter reading of
DTT and EGTA were from Sigma. Pipes was from Research
7.3. This change did not alter the results shown in Figures 1
The question of whether the effect of DMSO on the
products of assembly in D2O is reversible arose. To test this,
Assembly in D2O Buffer. In experiments to be described
tubulin was assembled in D2O/8% DMSO, and the products
later, we compare the assembly activity of tubulin after
were collected by centrifugation and suspended in H2O
incubation in buffer containing either H2O or D2O. To reduce
buffer. This protein was reassembled in H2O/8% DMSO at
the number of variables, the assembly reactions were carried
the same concentration as the original sample. Development
out under identical conditions, except for the presence or
of turbidity and the structure of the products were similar to
absence of D2O. Therefore, the reaction solutions contained
those that had not been preassembled in D2O/DMSO (Figures
8% DMSO. When the reactions were followed by turbidity
Biochemistry, Vol. 38, No. 10, 1999 3069
FIGURE 2: Structure of assembly products. Samples from the assembly reactions described in the legend of Figure 1 were negativelystained and examined by electron microscopy (40000× magnification) as described in Experimental Procedures. (A) Assembly in H2Obuffer with 8% DMSO. (B) Assembly in D2O buffer without DMSO. (C) Assembly in D2O buffer with 8% DMSO. (D) Assembly in D2Obuffer and then in H2O buffer, both containing 8% DMSO. Loss of Assembly ActiVity. Tubulin was incubated in buffer
over a reasonable time period, tubulin was incubated at 37
C. The results are presented in Figure 4. Although under
54 h. Samples were removed at different times, diluted to
these conditions the protein lost activity in D2O buffer, the
1.5 mg/mL, and tested for assembly competence. The amount
rate was much slower than in H2O buffer. In H2O buffer,
of polymerized tubulin was calculated from the protein
50% of the activity was lost in about 7 h, whereas in D2O
concentration in the cold-soluble pellet following centrifuga-
buffer, the activity had not decreased by 50% after 24 h.
tion and suspension of the pellet in cold PEM buffer. Figure
Exposure of Nonpolar Residues. BisANS is a probe for
3A depicts the results of these experiments. In H2O buffer,
nonpolar regions of proteins. Upon BisANS interacting with
tubulin had lost the ability to form microtubules within 40
hydrophobic regions, its fluorescence is greatly enhanced.
h. On the other hand, D2O afforded complete stability for
During the process of tubulin inactivation, the extent of
54 h. There was no significant amount of a cold-insoluble
binding of BisANS increases (9), indicating that hydrophobic
pellet formed during the assembly reaction in either H2O
regions become exposed. In Figure 5, data for the binding
of BisANS after various incubation periods in H2O buffer
The stability was also determined at 37 °C. In this case,
and D2O buffer are presented. D2O did decrease the rate at
tubulin was incubated for a period of 8 h. During this time,
which the hydrophobic regions are exposed to a small extent,
the sample in H2O buffer became turbid but the sample in
but the final percent change in fluorescence was equivalent
D2O buffer did not. After 8 h, the sample in H2O buffer had
in both solvents. Interestingly, the fluorescence value of
completely lost the ability to assemble, whereas the sample
BisANS and tubulin at zero time was 28-45% lower in D2O
in D2O buffer had retained 80% of its initial assembly activity
buffer than in H2O buffer, suggesting a lower degree of
binding in D2O. D2O had no effect on the fluorescence of
Samples incubated in both H2O buffer and D2O buffer for
8 h at 37 °C and 54 h at 4 °C were analyzed by SDS-
Aggregation of Tubulin. The observation that at 37 °C in
PAGE and showed no signs of degradation (data not shown).
H2O buffer the tubulin solution became turbid indicates that
Loss of Colchicine Binding. The colchicine binding activity
aggregation occurred. To determine whether aggregation also
of tubulin is lost at a slower rate than assembly activity (8).
occurred at 4 °C, size-exclusion HPLC was used. Freshly
To measure the decrease in the extent of colchicine binding
prepared tubulin showed one major elution peak at the elution
3070 Biochemistry, Vol. 38, No. 10, 1999
FIGURE 5: Increase in the extent of BisANS binding upon storage.
IGURE 3: Loss of assembly activity with time. (A) Tubulin at 2.2
Tubulin was incubated at 1.0 mg/mL in H2O and D2O buffer. At
2O buffer or D2O buffer was stored at 4 °C. At the
times indicated, samples were removed, diluted to 1.5 mg/mL, and
the times shown, samples were taken and diluted to 2 µM (0.2
made 8% in DMSO and 0.5 mM in GTP. The samples were
mg/mL) and BisANS was added to a final concentration of 10 µM.
assembled as described in Experimental Procedures. The concentra-
The fluorescence was read immediately using an excitation
tion of the cold-soluble pellet after centrifugation and suspension
wavelength of 385 nm and an emission wavelength of 490 nm.
in the original volume represents the concentration of polymerized
The band-passes were 2 nm each. The data shown came from a
tubulin formed. At time zero, this concentration was 1.1 mg/mL.
single experiment, and each data point is an average of duplicate
(B) Tubulin at 2 mg/mL was stored at 37 °C in H
readings. A second experiment gave similar results.
D2O buffer and treated as described for panel A. The concentrationof the cold-soluble pellet at time zero after suspension was 1.0 mg/mL.
FIGURE 4: Loss of colchicine binding activity upon storage. Tubulinwas incubated at 1.5 mg/mL and 37 °C in H2O buffer and D2Obuffer. At the times shown, samples were diluted to 5 µM (0.5mg/mL) and colchicine was added to a final concentration of 10
FIGURE 6: Increase in the level of tubulin aggregation upon storage. µM. The samples were incubated for 1 h at 37 °C, and the
Tubulin at 1 mg/mL was incubated at 4 °C in H2O buffer and D2O
fluorescence was read at 430 nm with an excitation wavelength of
buffer for the times shown. Samples were removed and injected
350 nm. The band-passes were 5 nm each. The values are
into a size-exclusion column as described in Experimental Proce-
normalized to 100% for the time zero value. The data shown
dures. (A) Elution patterns at 0 and 60 h. (B) The combined area
represent the average of two experiments. Both experiments gave
of the aggregate peak and the tubulin dimer peak was calculated,
a similar half-life of colchicine binding decay in H
and the percentage of the aggregate peak is plotted against time of
volume expected for a protein with a mass of 100 kDa
buffer, the aggregate peak represented only 16% of the total
(Figure 6A). A very small peak at the void volume was also
area (Figure 6A). The time course of the increase in the
noticeable. After 60 h at 4 °C in H2O buffer, the peak at the
amount of aggregate is presented in Figure 6B.
void volume had become the predominant one, representing
ReVersibility of InactiVation and Aggregation. The experi-
77% of the total area. In marked contrast, after 60 h in D2O
ments described in Figures 1 and 2 demonstrated that the
Biochemistry, Vol. 38, No. 10, 1999 3071
effect of D2O on the nature of the assembly product was
Studies of the effects of D2O on microtubules in vivo have
reversible. We then raised the question of whether the
been reported since 1935 (27). These studies have demon-
inactivation could be reversed by D2O. In an experiment in
strated that D2O increases the birefringence and volume of
which tubulin was stored for 36 h in H2O buffer at 4 °C, the
the mitotic spindle as well as the number and length of the
assembly activity had fallen to 19% of the original value.
spindle microtubules (reviewed in ref 28), indicating an
At this time, the protein was dialyzed against D2O buffer.
increase in the extent of tubulin polymerization caused by
After 4 h of dialysis, the activity increased to 50%. The H2O
D2O. D2O is also an antimitotic agent (29) and stabilizes
concentration of the dialysate was determined by 1H NMR
the spindle (30), suggesting that the deuterated solvent may
spectroscopy and was found to be 11%, the same as that of
affect the dynamics of mitotic microtubules. Stabilization
the dialysis buffer. Further dialysis did not restore any more
of microtubules may explain the antagonistic effect of D2O
activity. The products of assembly after dialysis were
on such antimitotic agents as colchicine (31) and methyl
examined in the electron microscope, and the structures were
benzimidazol-2-ylcarbamate (32). It has also been shown that
identical to those normally seen in assembly reactions carried
D2O stimulates tubulin assembly in vitro (33-35). The
out in D2O. The distribution between aggregate and dimer,
results presented in Figures 1 and 2 confirm these previous
which was 52% aggregate after the 36 h period, did not
findings. In the absence of DMSO and under the conditions
change after dialysis in D2O buffer. Thus, the amount of
used, assembly occurred in D2O but not in H2O.
reactivation was equivalent to the amount of dimer present.
Stimulation of tubulin assembly by D2O is most probably
Tubulin that had been incubated for 90 h and showed no
explained by the increase in the strength of hydrophobic
residual activity and little dimer present could not be
interactions in this solvent as proposed by Itoh and Sato (34)
because tubulin assembly is an entropically driven process(36). But D2O has a second effect on tubulin, preventing
DISCUSSION
inactivation and aggregation of the protein. Inhibition of theaggregation process is opposite to the effect of the solvent
A number of studies of the effects of D2O on protein
on tubulin polymerization and on the aggregation of the
aggregation have been reported. In many cases, D2O
proteins described above. It has been proposed that the
stabilized the aggregated form of oligomeric proteins. For
mechanism of tubulin inactivation involves a rate-limiting
example, the deuterated solvent stabilized the oligomeric
unimolecular conformational change before aggregation
form of halophilic malate dehydrogenase (10), lactic dehy-
occurs because the rate of tubulin aggregation does not show
drogenase (11), and glutamate dehydrogenase (11). D2O also
a dependence on protein concentration (1). This two-step
increased the level of aggregation of phycocyanin (12), the
process, which has also been proposed for the inactivation
level of association of the dimer form of -lactoglobulin A
of -lactoglobulin (22), is represented by the equation
to form the octamer (13), the rate of actin polymerization(14), the rate of polymerization of flagellin (15), and the
rate as well as the extent of subunit association of N10-formyl-tetrahydrofolate synthetase (16). Not all proteins are similarly
Undoubtedly, the process involves more than one intermedi-
affected by D2O though. For example, Berns et al. (12) found
ate step, and it is difficult to assign the step or steps that are
that the aggregation of several proteins was not affected by
affected by D2O. However, D2O would be expected to
the solvent. D2O is also known to affect the stability of some
decrease the rate of steps that involve conformational changes
proteins. The thermal stability of flagellin (15), ribonuclease
and the disruption of hydrophobic interactions in the dimer
(17), human factor XIII (18), and -lactoglobulin (19) and
because these interactions are stronger in D2O than in H2O.
the stability of ovalbumin in urea (20) are increased in D2O.
The experiments with BisANS showed that nonpolar regions
On the other hand, the thermal stability of val-tRNAval
are exposed during the inactivation process and the rate of
synthetase was unaffected by D2O (21).
exposure was decreased somewhat by D2O. But this effect
Despite the many studies, it has not been possible to
of D2O was not nearly as large as the effect on assembly
adequately describe the mechanism of the effects D2O on
activity and aggregation. Thus, exposure of nonpolar groups
protein structure (reviewed in ref 22). In a number of cases,
still occurs in D2O without the loss of activity or the
the effect of stabilizing the aggregated form of proteins has
production of aggregates. The BisANS binding data are
been attributed to the enhancement of hydrophobic inter-
difficult to assess, however, because it is not known what
actions because such interactions are stronger in D2O than
steps in the inactivation process contribute to the increase
in H2O (23). It has also been proposed that deuterium bonds
in the extent of BisANS binding and what effect aggregate
are stronger than hydrogen bonds (24), and several groups
formation has on the binding. It should also be pointed out
have suggested that D2O may have its effect, at least in part,
that in measuring inactivation, the protein was incubated at
on hydrogen bonding in proteins (13, 15, 20). However, in
0 °C but assembly was measured at 37 °C. We do not know
many cases in which the solvent isotope effect on hydrogen
whether the temperature shift has different consequences on
bonding was studied, enthalpic and entropic effects canceled
the protein in D2O and H2O. Very likely, D2O does not affect
each other and, as a result, there was little effect of D2O
the aggregation step because aggregation was not reversed
(7). Another factor that must be considered is the effect of
by the solvent, whereas the inactivation was reversed in
the solvent on the pKa values of amino acid residue side
proportion to the amount of dimer present in solution. It is
chains in the protein (25). The complexity of the solvent
more probable that D2O reversed a conformational(s) state
isotope effect can be seen in a study on the refolding of hen
to an active form(s) or perhaps prevented the inactivation
egg white lysozyme in which it was not possible to contribute
of a metastable state upon the temperature shift to 37 °C.
the effect to any one factor (26).
An interesting point is that D2O appeared to decrease the
3072 Biochemistry, Vol. 38, No. 10, 1999
extent of BisANS binding to tubulin because the fluorescence
Ed.) Advances in Chemistry Series, pp 21-30, American
that tubulin has a more compact structure in D
13. Baghurst, P. A., Nichol, L. W., and Sawyer, W. H. (1972) J.
It has been proposed that the tubulin aggregation process
14. Omori, H., Kuroda, M., Naora, H., Takeda, H., Nio, Y., Otani,
involves the formation of disulfide bonds (1). Our studies
H., and Tamura, K. (1997) Eur. J. Cell Biol. 74, 273-280.
were carried out in the presence of the reducing agent DTT
15. Uratani, Y. (1974) J. Biochem. (Tokyo) 75, 1143-1145.
which should have prevented disulfide formation. Incubation
16. Harmony, J. A. K., Himes, R. H., and Schowen, R. L. (1975)
of tubulin at 37 °C for periods of up to 30 days also results
in the formation of isoaspartyl bonds (37) and incubation at
17. Hermans, J., Jr., and Sheraga, H. A. (1959) Biochim. Biophys.
high pH and temperature in lysinoalanine cross-links (38),
18. Dong, A., Kendrick, B., Kreigård, L., Matsuura, J., Manning,
but these processes probably do not contribute to the changes
M. C., and Carpenter, J. F. (1997) Arch. Biochem. Biophys.
we observed over a much shorter time period at 4 °C and
19. Verheul, M., Roefs, S. P. F. M., and de Kruif, K. G. (1998)
Why are the structures formed during assembly in D2O/
20. Maybury, R. H., and Katz, J. J. (1956) Nature 177, 629-630.
21. Kern, D., Zaccaı´, G., and Giege, R. (1980) Biochemistry 19,
and DMSO are stimulators of tubulin assembly,
lowering the critical protein concentration for polymerization.
22. Oas, T. G., and Toone, E. J. (1997) AdV. Biophys. Chem. 6,
There is possibly more than one pathway to polymerization.
One pathway leads to nuclei that form microtubules, and
23. Kresheck, G. C., Schneider, H., and Scheraga, H. A. (1965)
another pathway might lead to nuclei that are seeds for ribbon
J. Phys. Chem. 69, 3132-3144.
formation. The combination of D2O and DMSO could
24. Ne´methy, G., and Sheraga, H. A. (1964) J. Chem. Phys. 41,
increase the rate of formation of nuclei that lead to ribbons.
This is similar to what happens when the DMSO concentra-
25. Bunde, A, and Wu¨thrich, K. (1979) Biopolymers 18, 299-
tion in H2O is above 10% (39) or high concentrations of
26. Itzhaki, L. S., and Evans, P. A. (1996) Protein Sci. 5, 140-
organic sulfonate salts are present (40).
27. Lucke´, B., and Harvey, E. N. (1935) J. Cell. Comp. Physiol.ACKNOWLEDGMENT
We thank Dr. David Vander Velde, Director of the
28. Sato, H., Kato, T., Takahashi, T. C., and Ito, T. (1982) in
Biological Functions of Microtubules and Related Structures
University of Kansas NMR laboratory, for conducting the
(Sakai, H., Mohri, H., and Borisy, G. G., Eds.) pp 211-226,
analysis of the H2O content of D2O solutions.
29. Marsland, D. A., and Zimmerman, A. M. (1963) Exp. CellREFERENCES
30. Marsland, D., and Zimmerman, A. M. (1965) Exp. Cell Res.
1. Prakash, V., and Timasheff, S. N. (1982) J. Mol. Biol. 160,
31. Marsland, D., and Hecht, R. (1968) Exp. Cell Res. 51, 602-
2. Tiwari, S. C., and Suprenant, K. A. (1993) Anal. Biochem.
3. Algaier, J., and Himes, R. H. (1988) Biochim. Biophys. Acta
32. Howard, R. J., and Aist, J. R. (1980) J. Cell Biol. 87, 55-64.
33. Olmsted, J. B., and Borisy, G. G. (1973) Biochemistry 12,
4. Himes, R. H., Burton, P. R., and Gaito, J. M. (1977) J. Biol.
34. Houston, L. L., Odell, J., Lee, Y. C., and Himes, R. H. (1974)
5. Bradford, M. M. (1976) Anal. Biochem. 72, 248-256.
6. Bhattacharyya, B., and Wolff, J. (1974) Proc. Natl. Acad. Sci.
35. Ito, T., and Sato, H. (1984) Biochim. Biophys. Acta 800, 21-
7. Schowen, B. K., and Schowen, R. L. (1982) Methods Enzymol.
36. Correia, J. J., and Williams, R. C., Jr. (1983) Annu. ReV.Biophys. Bioeng. 12, 211-235.
8. Wiche, G., Honig, L. S., and Cole, R. D. (1977) Nature 269,
37. Najbauer, J., Orpiszewski, J., and Aswad, D. W. (1996)
9. Prasad, A. R. S., Luduen˜a, R. F., and Horowitz, P. M. (1986)
38. Correia, J. J., Lipscomb, L. D., and Lobert, S. (1993) Arch.Biochem. Biophys. 300, 105-114.
10. Bonnete, F., Madern, D., and Zaccai, G. (1994) J. Mol. Biol.
39. Himes, R. H., Burton, P. R., Kersey, R. D., and Pierson, G.
B. (1976) Proc. Natl. Acad. Sci. U.S.A. 73, 4397-4399.
11. Henderson, R. F., Henderson, T. R., and Woodfin, B. H. (1970)
40. Himes, R. H., Newhouse, C. S., Haskins, K. M., and Burton,
J. Biol. Chem. 245, 3733-3737.
P. R. (1979) Biochem. Biophys. Res. Commun. 87, 1031-1038.
12. Berns, D. S., Lee, J. J., and Scott, E. (1968) in MolecularAssociation in Biological and Related Systems (Gould, R. F.,
■ Chimiothérapie et dans les cancers ORL, alors que le libelléLimousin, ont été mises en œuvre en 2002 des cancers pulmonaires : de l’AMM le réserve aux cancers du sein etpour notamment améliorer les conditions de un état des lieux préoccupant préparation des chimiothérapies et mieuxinformer les établissements des démarches Dans une dépêche, l’agence APM-Reuters,
Referência : http://www.time.com/time/time100/scientist/profile/fleming.html Alexander Fleming A spore that drifted into his lab and took root on a culture dish started a chain of events that altered forever theThe improbable chain of events that led Alexanderbacteriologist." Although he went on to performFleming to discover penicillin in 1928 is the stuff ofadditional experiments, h
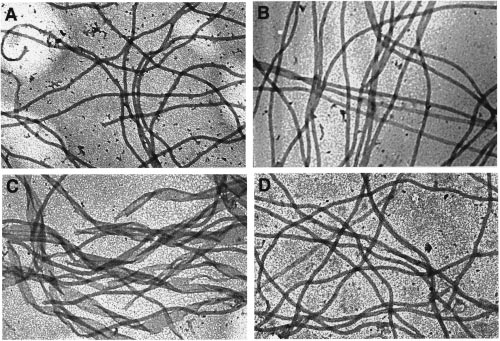 Biochemistry, Vol. 38, No. 10, 1999 3069
FIGURE 2: Structure of assembly products. Samples from the assembly reactions described in the legend of Figure 1 were negativelystained and examined by electron microscopy (40000× magnification) as described in Experimental Procedures. (A) Assembly in H2Obuffer with 8% DMSO. (B) Assembly in D2O buffer without DMSO. (C) Assembly in D2O buffer with 8% DMSO. (D) Assembly in D2Obuffer and then in H2O buffer, both containing 8% DMSO.
Biochemistry, Vol. 38, No. 10, 1999 3069
FIGURE 2: Structure of assembly products. Samples from the assembly reactions described in the legend of Figure 1 were negativelystained and examined by electron microscopy (40000× magnification) as described in Experimental Procedures. (A) Assembly in H2Obuffer with 8% DMSO. (B) Assembly in D2O buffer without DMSO. (C) Assembly in D2O buffer with 8% DMSO. (D) Assembly in D2Obuffer and then in H2O buffer, both containing 8% DMSO.