Kamagra enthält Sildenafilcitrat als pharmakologisch aktiven Bestandteil. Dieser hemmt selektiv die Phosphodiesterase-5 und erhöht dadurch die Konzentration von cGMP im Corpus cavernosum. Der Effekt ist zeitlich begrenzt, da die Halbwertszeit von Sildenafil etwa vier Stunden beträgt. In der galenischen Form als Mundgel erfolgt die Resorption besonders rasch, was zu einem schnelleren Wirkeintritt führt. Der Abbau erfolgt überwiegend hepatisch über CYP3A4, wobei ein aktiver Metabolit entsteht, der zur Gesamtwirkung beiträgt. Typische Nebenwirkungen ergeben sich aus der Vasodilatation, darunter leichte Kopfschmerzen und nasale Kongestion. In klinischen Beschreibungen wird kamagra oral jelly im Zusammenhang mit der schnelleren Absorption erwähnt.
Ortobrandao.com.br
Psychopharmacology (2002) 159:138–144DOI 10.1007/s002130100883
Luiz Carlos Schenberg · Larissa Bustamante Capucho Ricardo Ossamu Vatanabe · Leila César Vargas
Acute effects of clomipramine and fluoxetine on dorsal periaqueductalgrey-evoked unconditioned defensive behaviours of the rat
Received: 6 December 2000 / Accepted: 7 July 2001 / Published online: 22 September 2001 Springer-Verlag 2001
Abstract Rationale: Several antidepressants attenuate Keywords Clomipramine · Fluoxetine · 5-HT · Panic ·
conditioned escape behaviours reinforced by the terminus
Periaqueductal Grey · Freezing · Flight
of an electrical stimulus applied to the dorsal periaque-ductal grey (DPAG). Objective: The present study exam-ined whether the antidepressant and antipanic drugs
clomipramine (CLM) and fluoxetine (FLX) also attenuatethe DPAG-evoked unconditioned defensive behaviours.
Electrical stimulation of dorsal periaqueductal grey
Methods: Rats with electrodes in the DPAG were
matter (DPAG) has putative aversive properties because
electrically stimulated in the absence of any treatment or
rats readily learn to avoid or switch-off the stimulus.
30 min after injections of CLM, FLX or saline. Threshold
Electrical and chemical stimulations of DPAG also produce
functions of cumulative response frequencies were fitted
unconditioned defensive behaviours. Thus, either a
through the logistic model and compared using likelihood
freezing behaviour characterised by a tense immobile
ratio coincidence tests. Results: CLM produced non-
display, exophthalmus, vibrissae paralysis and/or defecation
linear effects on galloping, for which median thresholds
and micturition, or a flight behaviour made up of
(I ) were significantly increased (19±2%) or decreased
trotting, galloping and jumping responses is brought
(–22±2%) with 5 mg/kg and 10 mg/kg, respectively, or
about by electrical or chemical stimulation of DPAG
did not change with 20 mg/kg. The latter dose further
(Sudré et al. 1993; Bittencourt et al. 2000; Schenberg et
al. 2000; Vargas et al. 2000). Although the stimulation of
the defecation output (–33±15%). FLX significantly
DPAG has been formerly proposed as an anxiety model
in operant procedures (Schenberg and Graeff 1978;
(25±3%) with 1 mg/kg and 5 mg/kg, respectively. More-
Graeff 1981), DPAG stimulation in healthy humans
over, corresponding doses either decreased the maximum
produces sensations, visceral responses and neurological
symptoms remarkably similar to clinical panic attacks
defecation. Saline was ineffective. Conclusions: While
(Nashold et al. 1969). Therefore, DPAG stimulation has
the attenuation of defecation and micturition by
also been proposed as a model of panic attacks (Gentil
20 mg/kg CLM suggests a peripheral antimuscarinic action,
1988; Deakin and Graeff 1991). As a matter of fact,
CLM non-linear effects on galloping were most likely
pharmacological evidence in the rat supports the
due to its differential action on monoaminergic and
panic-like nature of DPAG stimulation. Thus, while the
cholinergic central mechanisms. In contrast, the attenuation
acute administration of drugs that are effective in panic
of immobility, galloping and defecation by low doses of
therapy (panicolytics) – such as the selective serotonin
FLX suggests a serotonin-mediated antiaversive action.
(5-HT) re-uptake inhibitors (SSRIs), fluoxetine (FLX),
Finally, CLM and FLX acute effects on DPAG-evoked
fluvoxamine and sertraline or the high-potency benzodi-
unconditioned galloping response were strikingly
azepines alprazolam and clonazepam – attenuated the
similar to those reported for DPAG-evoked shuttle-box
DPAG-evoked shuttle-box conditioned escape, drugs that
are known to precipitate panic attacks (panicogenics) –such as yohimbine, caffeine and cholecystokinin receptor
L.C. Schenberg (✉) · L.B. Capucho · R.O. Vatanabe · L.C. Vargas
agonists – had a facilitatory effect (Jenck et al. 1990,
Departamento de Ciências Fisiológicas – Centro Biomédico,
1995, 1996, 1998). However, the clinically effective
panicolytics imipramine and clomipramine (CLM) were
Av. Marechal Campos 1468 (Maruípe), 29040-090 Vitória-ES, Brazil
ineffective in the DPAG-evoked shuttle-box escape
e-mail: schenber@terra.com.brTel.: +55-27-33357332, Fax: +55-27-33357330
(Jenck et al. 1990). In contrast, recent studies from our
laboratory showed that the administration of CLM at a
time course and dose regimen (5 mg/kg and 10 mg/kg/day,
Only rats that exhibited jumps with peak-to-peak intensities below
21 days) not much different from those observed in panic
55 µA were selected for drug treatments. CLM (clomipramine
therapy significantly increased the thresholds of DPAG-
hydrochloride, Sigma, St. Louis, Mo.) and FLX (fluoxetine hydro-
evoked immobility (24%), trotting (138%), galloping
chloride, Eli Lilly, São Paulo, Brazil) were dissolved in saline
(75%), jumping (45%) and micturition (87%). In addition,
(0.9% NaCl) and distilled water, respectively. CLM (5, 10 and
the 21-day administration of FLX (1 mg/kg/day) virtually
20 mg/kg, i.p., n=28, 18 and 18, respectively) and FLX (1 mg/kgand 5 mg/kg, i.p., n=20 and 22, respectively) were administered
abolished galloping without changing the remaining
30 min before the onset of stimulation sessions. Controls were
responses (Vargas and Schenberg 2001).
similarly treated with saline (n=24). Drug sedative effects on
There are no studies, however, evaluating the acute
open-field ambulatory activity were evaluated in separate rats
effects of panicolytics on unconditioned defensive
(n=10 per group) similarly treated with saline, FLX (1 mg/kg and5 mg/kg) or CLM (5 mg/kg and 10 mg/kg).
behaviours produced by electrical stimulation of DPAG. Accordingly, the present study employed the thresholdlogistic analysis (Sudré et al. 1993; Schenberg et al.
1990, 2000) to assess the acute effects of the clinically
The rat behavioural output was rated according to a previously
effective panicolytics CLM and FLX on DPAG-evoked
made ethogram (Schenberg et al. 2000). The following responses
were recorded: sleeping, resting, grooming, rearing, walking, tenseimmobility, trotting, galloping, jumping, exophthalmus (eyeballprotrusion and wide opened eyelids), defecation and micturition.
Furthermore, vibrissae paralysis (mystacioplegia) was alsoobserved during the freezing behaviour. Behaviours were recordedin a binary way, as emitted or not, irrespective of their frequency
or duration in a single-stimulation trial. In order to determine theresponse threshold curves, only the “threshold responses”, i.e.
Male albino Wistar rats (200–250 g) were housed in individual
those emitted with the minimally effective current, were subjected
glass-walled cages with food and water ad libitum and a natural
to statistical analysis. Trotting and galloping were analysed either
separately or merged as the “running” response. Moreover,because defecation and micturition also occur spontaneously,unbiased fitting of their stimulus-dependent output was performed,
discarding the responses emitted during 0- (sham), 5- and 10-µAstimulation trials, supposedly, due to the rat exploratory activity.
Rats were anaesthetised with 400 mg/kg (i.p.) chloral hydrate (Isofar,Rio de Janeiro, Brazil) supplemented by the s.c. infiltration ofscalp with 1% lidocaine plus 0.005% epinephrine (Cristália, São
Paulo, Brazil). Thereafter, rats were fixed on a stereotaxic apparatus(David Kopf, Tujunga, Calif.) and wrapped with a cloth to avoid
Drug sedative effects were assessed in separate groups by means
surgical hypothermia. With the skull horizontal between bregma
of a custom-built infrared-based actometer. The actometer was
and lambda, the bone over the lambda was abraded with the aid of
made up of an anti-burglary domestic system (Infraset, São Paulo,
a drill and removed with thin forceps to expose the sinus.
Brazil) in which the time-constant circuit was modified to allow a
Monopolar stainless-steel electrodes (o.d. 200 µm), insulated
faster resetting. The actometer had a 360° monitoring radius and
throughout except at the cross-section of the tip, were then inserted
was placed 50 cm above the floor of the open field. The sensor
in the right or left dorsal midbrain through a small dura incision
was sensitive to a 12-cm mean displacement of the rat and filtered
just by the sinus. Whenever necessary, the sinus was gently
slow stretching responses, scratching, grooming and small move-
pushed with the electrode itself so as to allow its penetration to the
ments of the paws and head. Accordingly, records denote the
aimed site. The electrode was anchored to the skull by means of a
ambulatory activity only (walking and rearing). Open-field
U-shaped, stainless-steel clip, three small screws and dental resin.
sessions were carried out between 0800 hours and 1500 hours in a
In addition, the bone was covered with a thin layer of cyanoacrylate
sound-attenuated, temperature-controlled room (23–25°C). Naive
methylester glue (Super-Bond, São Paulo, Brazil) just before
rats were injected (i.p) with saline, FLX or CLM and placed after
pouring the dental resin to hold the pieces together. These procedures
30 min into the open field. Cumulative ambulatory scores of 10-,
aided in keeping the electrodes in place for a 30-day period or so
20- and 30-min recording periods (i.e., 40, 50 and 60 min after
allowing the chronic treatment with CLM and FLX reported
drug injection) were subjected to statistical analysis.
elsewhere (Vargas and Schenberg 2001).
At the end of experiments, brains were sectioned in a freezing
Rats were stimulated in a cylindrical Plexiglass open-field apparatus
microtome (60-µm sections) and stained with neutral red. Stimulation
of 60-cm wall height and diameter. The screening sessions
sites were plotted on diagrams of the rat brain atlas of Paxinos and
were carried out 4–10 days after surgery. The rats were connected
to a constant current sine-wave stimulator and placed into theopen field, where they remained undisturbed for 15 min to getused to the environment and reduce spontaneous activity. Following
this period, stepwise increasing stimuli (0–55 µA, 60 Hz,1 min, a.c.) were presented at 5-min intervals. A light cable and
Behavioural items were recorded in a binary way, as emitted or
mercury swivel allowed the free movement of the rat during brain
not, irrespective of their frequency or duration in a single stimulation
stimulation. In each trial, the intensity was increased by 5 µA until
trial. Response threshold curves were obtained by logistic fitting
the rat showed the jumping response. All experiments were
of accumulated response frequencies. Significant regression on
carried out in a sound-attenuated, temperature-controlled room
stimulus intensity was assessed using Wald’s chi-square test. Drug
effects were assessed according to a within-subject design. Pre-
and post-drug threshold curves of each group were modelled bymeans of indicator variables and compared for either location(triggering level) or parallelism (responsiveness) using likelihood-ratio coincidence tests. Behaviour triggering level was representedby estimates of median intensity (I ±SE). Behavioural respon-
siveness was represented by regression curvature (β±SE), i.e. theparameter that governs the rate of change in response probabilityas a function of stimulus change. Finally, whenever a low responseoutput precluded the logistic fitting, maximum accumulatedfrequencies (P
using odds ratio (ψ) and Pearson’s χ2 analyses. A comprehensivedescription of our original approach to the threshold logistic analysisof intracranially induced behaviours can be found elsewhere(Schenberg et al. 2000).
Drug sedative effects on open-field ambulatory behaviour were
evaluated using repeated-measures analysis of variance (ANOVA)followed by planned contrasts (1 d.f.) for time × group interactions. Pairwise multiple comparisons were considered significant atBonferroni’s 5% level. All statistical analyses were performedusing the SAS software (Statistical Analysis Systems, N.C.).
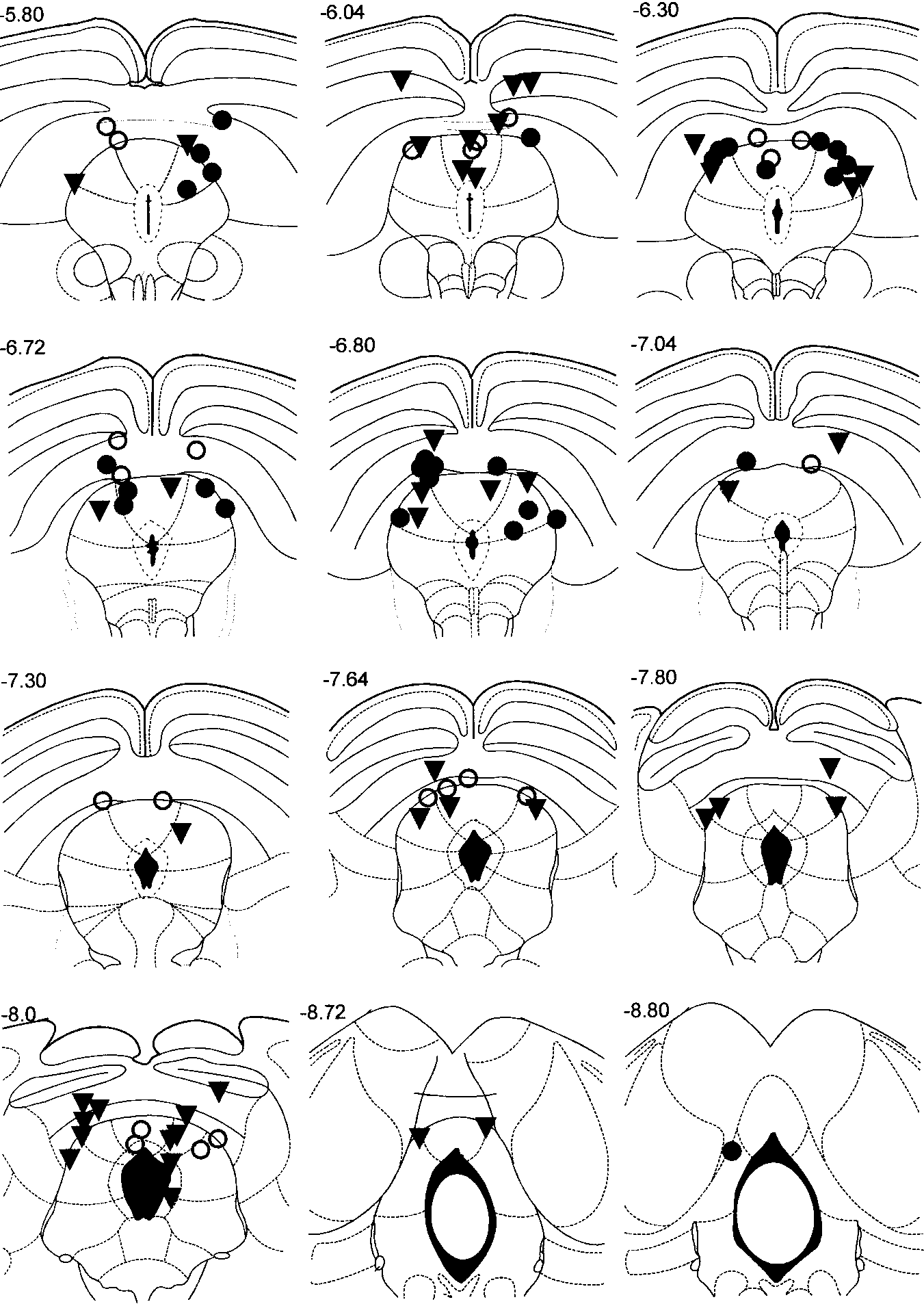
Histology was performed in 96 of 130 rats. The remaining ratswere lost throughout the ensuing long-term treatment with CLMand FLX (Vargas and Schenberg 2001). Electrodes within thedorsomedial and dorsolateral columns of DPAG or just borderingit comprised 78%. The remaining electrodes were localised in thedeep collicular layers (intermediate layer 7%, deep grey/whitelayers 15%). According to the current parcellation of the PAG (P. Carrive, personal communication), most sites were distributedthroughout the intermediate (53) and caudal (29) DPAG or adjoiningdeep collicular layer (bregma AP coordinates: –6.04 mm to–7.3 mm). Moreover, seven electrodes were localised in the rostralDPAG (–5.8 mm; Fig. 1). Stimulation of these sites yielded signifi-
Fig. 1 Brain sites in which stimulation during pre-drug sessions
cant regressions for defensive responses only. Pooled pre-drug
elicited the jumping response with intensities below 55 µA (60 Hz,
sessions (n=130) showed the following threshold hierarchy:
1 min, a.c.). Plates represent the coronal sections of the rat brain
immobility < exophthalmus < jumping < trotting < galloping <
atlas of Paxinos and Watson (1986). Numbers are the rostro-
micturition < defecation (Table 1). Distribution of sites within the
caudal coordinates in relation to bregma (mm). Clomipramine
DPAG and deep collicular layers did not differ among drug groups
(filled inverted triangle), fluoxetine (filled circle) and saline (open
Table 1 Rat defensive profile of dorsal periaqueductal gray
CLM produced non-linear effects on galloping, for which the
stimulation in pooled pre-drug sessions (n=130). I
thresholds were either increased (∆I =19±1.8%, χ2=7.5, d.f.=1,
(the estimate of triggering level, i.e. the intensity in which a given
P<0.006), decreased (∆I =–22±2.1%, χ2=6.1, d.f.=1, P<0.01) or
response has the higher frequency), β curvature parameter (the
did not change with 5, 10 and 20 mg/kg, respectively (Fig. 2).
estimate of responsiveness, i.e. the rate of change in response
Thresholds of micturition were increased by 20 mg/kg CLM
probability in function of stimulus change), P
(∆I =38±1.1%, χ2=3.6, d.f.=1, P<0.05). This dose also reduced
response output (the estimate of stimulus efficacy, i.e. the maximum
the maximum output of defecation. Indeed, the defecation pre-
accumulated frequency). Responses were sorted according to their
drug probability was six times higher than that of rats treated with
20 mg/kg CLM (ψ=6.4±5.6, χ2=4.9, d.f.=1, P<0.02). Immobility,exophthalmus, trotting, running and jumping responses did not
FLX produced significant increases in the thresholds of immobility
(∆I =22±2.1%, χ2=9.4, d.f.=1, P<0.002) and galloping
(∆I =25±2.6%, χ2=12.0, d.f.=1, P<0.0005) with 1 mg/kg and
5 mg/kg, respectively (Fig. 3). Defecation was also attenuated by
1 mg/kg and 5 mg/kg FLX which caused a fivefold decrease inoutput probability (ψ=4.8±4.2, χ2=3.6, d.f.=1, P<0.05) or asignificant increase in I
P<0.03), respectively. No changes were observed in exophthalmus,trotting, running, micturition and jumping responses. Fig. 2 Acute effects of clomipramine (CLM 5, 10 and 20 mg/kg, i.p.) and saline (SAL 0.9% NaCl, i.p.) on the thresh- olds and maximum output of galloping, defecation and micturition evoked by electrical stimulation of dorsal periaque- ductal grey. Sigmoidal curves represent the best-fitting logistic function of accumulated response frequencies. Line-plus-symbol graphs represent data that did not achieve a significant logistic fitting (r responders, n number of stimulated rats). Dashed curves (–-) and asterisks (*), P<0.05, represent curve location and maximum output signifi- cantly different from pre-drug controls, respectively (likelihood ratio coincident tests and maximum output Pearson’s χ2 as appropriate)
(10 mg/kg) facilitation of galloping may be associated with theintensification of anxiety and panic attacks seen in treatment onset
Whatever the regression parameter (I , β, P
(Ramos et al. 1993). The neurochemical mechanisms underlying
of saline did not change any DPAG-evoked defensive behaviour.
this “first-dose effect” remain obscure. However, besides their5-HT and noradrenaline reuptake inhibitory actions, tricyclic anti-depressants have long been known for their muscarinic blocking
activity and respective side effects, such as constipation, urinaryretention, blurred vision and dry mouth (Atkinson and Ladinsky
Compared with pre-drug controls, CLM-, FLX- and saline-treated
1971; Richelson and Divinetz-Romero 1977; Snyder and Yamamura
rats showed a reduced open-field activity in drug sessions carried
1977; Blackwell et al. 1978; Cusack et al. 1994). Moreover, rather
out the day after (Fig. 4). Nevertheless, while the open-field activity
than peripherally, imipramine seems to exert its anti-enuretic
of rats treated with 5 mg/kg CLM or 1 mg/kg and 5 mg/kg FLX
effect via a supraspinal antimuscarinic action (Sohn and Kim
did not differ from saline-treated ones, 10 mg/kg CLM significantly
1997). Thus, CLM non-linear effects on galloping were most
reduced the ambulatory activity at 20-min and 30-min recording
probably due to drug or metabolite differential effects on
serotonergic, adrenergic and cholinergic synapses. Indeed, it has
long been reported that scopolamine has a pro-aversive effect,
further decreasing the operant lever pressing which was suppressedby punishment (Miczek 1973).
In contrast to the complete absence of effects of saline, FLX
caused modest but significant attenuation of DPAG-evoked defensivebehaviours. The attenuation of galloping by a non-sedative dose of
Compared with chronically treated rats (Vargas and Schenberg
FLX (5 mg/kg) supports the 5-HT inhibitory modulation of flight.
2001), the acute administration of CLM produced fewer and lesser
Indeed, iontophoretic application of 5-HT predominantly inhibited
effects on DPAG-evoked defensive behaviours. Hence, while
single neuron activity in the dorsolateral and lateral sectors of
galloping was attenuated by a non-sedative dose of CLM
PAG (Lovick 1994). Moreover, these inhibitory actions were
(5 mg/kg), this response was facilitated following the administration
potentiated by the iontophoretic application of a SSRI panicolytic,
of a manifestly depressant dose (10 mg/kg). Accordingly, CLM
paroxetine, supporting the FLX inhibitory modulation of PAG
attenuation of galloping did not correlate with its sedative properties,
(Lovick 1994). Because galloping was attenuated to a much greater
supporting a specific anti-aversive action instead. However, CLM
extent following a 21-day administration of 1 mg/kg/day FLX
Fig. 4 Fluoxetine (FLX 1 mg/kg and 5 mg/kg, i.p.) and clomipramine (CLM 5 mg/kg and 10 mg/kg, i.p.) acute effects on open-field exploratory activity. Pre-drug sessions were performed the day Fig. 3 Acute effects of fluoxetine (FLX 1 mg/kg and 5 mg/kg, i.p.)
before drug testing. Activity was recorded 10, 20 and 30 min after
and saline (SAL 0.9% NaCl, i.p.) on the thresholds and maximum
session onset. *P<0.05, significantly different from saline-treated
output of immobility, galloping and defecation evoked by electrical
rats (repeated-measures analysis of variance followed by planned
stimulation of dorsal periaqueductal grey. Sigmoidal curves
contrasts for time × group interactions)
represent the best-fitting logistic function of accumulated responsefrequencies. Line-plus-symbol graphs represent data that did notachieve a significant logistic fitting (r responders, n number of
It is worth noting that exogenous 5-HT sensitises normal peristalsis
stimulated rats). Dashed curves (---) and asterisks (*), P<0.05,
and facilitates stress- and corticotropin releasing factor (CRF)-
represent curve location and maximum output significantly different
from pre-drug controls, respectively (likelihood ratio coincident
1992, 1998; Sanger et al. 1998). Accordingly, 5-HT has been
tests and maximum output Pearson’s χ2 as appropriate)
proposed as a key sensitising agent in the aetiology of irritablebowel syndrome, a functional gastrointestinal disorder (Sanger1996). In addition, central 5-HT
(Vargas and Schenberg 2001), both short- and long-term mechanisms
normal peristalsis (Croci et al. 1995). Consequently, FLX attenuation
seem to be involved in FLX inhibitory effects on galloping.
of DPAG-evoked defecation was most probably a specific anti-
Alternatively, galloping attenuation could have been a by-product
aversive action at DPAG properly or its efferent pathways. Indeed,
of a facilitation of freezing brought about by the enhancement of
recent transneuronal track tracing studies with pseudorabies virus
5-HT transmission in the “behavioural inhibition system” (Gray
suggested that colonic motility is chiefly controlled by Barrington’s
1991). Yet, instead of a facilitation, immobility was attenuated or
nucleus, formerly considered solely as a micturition centre, but
did not change after acute administration of 1 mg/kg and 5 mg/kg
also by the lateral PAG and spinally projecting locus coeruleus
FLX, respectively. Therefore, 5-HT seems to inhibit both freezing
CRF neurons (Monnikes et al. 1994; Valentino et al. 2000). In
and flight behaviours. Immobility attenuation, however, wanes
turn, the PAG is known to project to Barrington’s nucleus and
following the chronic administration of FLX (Vargas and Schenberg
locus coeruleus rostromedial dendrites (Valentino et al. 1994;
2001). Finally, attenuation of galloping but not immobility
Luppi et al. 1995; Blok and Holstege 1996). These data give neuro-
following the administration of 5 mg/kg FLX suggests a complex
anatomical support to the elicitation of defecation and micturition
interplay of DPAG-evoked somatic defensive behaviours. Apparently,
by electrical and chemical stimulation of PAG (Schenberg et al.
galloping attenuation with the higher dose of FLX shifted the
1990, 2000; Sudré et al. 1993; Bittencourt et al. 2000; Vargas et al.
defensive repertoire towards immobility and trotting, compensating
2000; Vargas and Schenberg 2001). Further, 5-HT inhibits the
an eventual attenuation of the latter responses. As a matter of fact,
PAG and locus coeruleus neuron activity, supporting the FLX
a similar interplay of galloping and trotting was also observed
inhibitory modulation of DPAG-evoked defecation (Shiekhattar
following the chronic treatment with FLX (Vargas and Schenberg
and Aston-Jones 1993; Lovick 1994). More importantly, given the
2001). In any event, FLX seems to downgrade the defensive
high co-morbidity of panic disorder and irritable bowel syndrome
repertoire from flight to freezing and thence to non-defensive
(Lydiard et al. 1994; Lydiard and Falsetti 1999), FLX attenuation
of DPAG-evoked defecation is likely to underlie the reported
Remarkably, whereas the attenuation of defecation and micturi-
effectiveness of antipanic agents on concomitant panic and irritable
tion by 20 mg/kg CLM could be due to a peripheral cholinergic
bowel disorders (Lydiard et al. 1986).
blockade, DPAG-evoked defecation was also attenuated by FLX,
Antidepressant effects confirmed previous studies with DPAG-
which is devoid of any antimuscarinic action (Cusack et al. 1994).
evoked shuttle-box escape behaviour. In particular, while the shuttle-
box escape was attenuated by SSRIs, FLX (10–32 mg/kg), sertraline
tasks such as bar-pressing or shuttle-box escape. Finally, the threshold
(1–10 mg/kg) and fluvoxamine (10–22 mg/kg), no threshold
logistic analysis detected effects of CLM and FLX doses within the
changes were observed following 10-mg/kg and 22-mg/kg doses
true clinical range. In this regard, it should be stressed that the
of the non-selective tricyclic antidepressants imipramine and CLM
thresholds of jumping of the present study were lower than those of
(Jenck et al. 1990). Likewise, in the present study, while galloping
trotting and galloping. The stringent criterion employed in rat
was attenuated by a non-sedative dose of FLX (5 mg/kg), it was
screening for drug treatments, i.e. selection of rats that jumped with
facilitated or did not change in the same dose range of CLM that
less than 55 µA, most probably biased the sample defensive profile
failed in attenuating the DPAG-evoked shuttle-box escape.
towards the high-threshold, drug-resistant jumping behaviour.
Notably, CLM had no effects on running (merging of trotting and
Indeed, immobility, trotting, galloping and jumping are usually
galloping), the behaviour most likely recorded in the shuttle-box
elicited with increasing stimuli when rats are pooled on either a
escape paradigm. The striking similarity of antidepressant actions
plain neuroanatomical basis (Bittencourt et al. 2000) or through a
on innate and shuttle-box escape behaviours thus supports the
more flexible criterion, i.e. presentation of any flight response, such
unconditioned nature of the latter behaviour. Consequently,
as trotting, galloping or jumping, with less than 50 µA (Sudré et al.
instead of a “conditioning”, DPAG-evoked shuttle-box escape
1993). Thus, the rather stringent criterion of rat selection should be
would be more akin to a stimulus titration procedure.
considered in the appraisal of present drug effects.
The neurochemistry of DPAG-evoked innate behaviours and
In conclusion, in contrast to the CLM non-linear effects on
their complex interplay remains unclear. Indeed, most studies on
galloping, FLX attenuation of immobility and galloping with
this issue were carried out with the shuttle-box escape procedure
non-sedative doses suggests a 5-HT antiaversive action similar to
and provide information on running responses only. However, as
that reported for FLX and other SSRIs on DPAG-evoked shuttle-box
far as the 5-HT transmission is concerned, it is noteworthy that
escape behaviour. Moreover, while the attenuation of defecation
intra-periaqueductal injection of both 5-HT and the SSRI zimelidine
and micturition by the higher dose of CLM suggests a peripheral
produced antiaversive effects (Schütz et al. 1985). Administration
antimuscarinic effect, FLX attenuation of defecation supports a
of 5-HT receptor agonists and antagonists yielded, however,
conflicting results. Therefore, while the systemic injection of thesedrugs suggested a pro-aversive role of both 5-HT
Acknowledgements This work was supported by research grants
receptor subtypes, the local injection of corresponding drugs led to
from AFIP, CNPq, FACITEC and FINEP. L.C. Schenberg, L.C.
opposite results (Jenck et al. 1989; Nogueira and Graeff 1995).
Vargas and L.B. Capucho were recipients of Research, Master
Thus, either the loss of drug specificity at high concentrations
Degree and Undergraduate CNPq fellowships, respectively. The
attained by intracerebral injections or the different sites of action
authors wish to thank Eli Lilly do Brasil Ltd. (São Paulo, SP) for
of systemically and locally injected drugs could underlie these
conflicting data. Indeed, DPAG-evoked unconditioned flightin the open-field was either enhanced or attenuated by the5-HT
selective agonist, 8-hydroxy-2-(di-n-propylamino)-tetralin
(8-OHDPAT), depending on the route of administration, systemic
or local, respectively (Beckett and Mardsen 1997).
It is also noteworthy that the predator-elicited flight behaviour
Atkinson J, Ladinsky H (1972) A quantitative study of the
was potentiated by acute administration of imipramine (5, 10 and
anticholinergic action of several tricyclic antidepressants on
15 mg/kg), which either reduced the avoidance distance or
the rat isolated fundal strip. Br J Pharmacol 45:519–524
increased the flight speed (Griebel et al. 1995, 1996; Blanchard et
Beckett S, Marsden CA (1997) The effect of central and systemic
al. 1997). A similar effect on galloping was observed in the present
study after the administration of a 10-mg/kg dose of CLM, which
receptor antagonist WAY100635 on periaqueductal
both reduced the triggering level and increased the responsiveness
grey-induced defence behaviour. J Psychopharmacol 11:35–40
(parameters akin to “avoidance distance” and “flight speed”,
Bittencourt AS, Carobrez AP, Schenberg LC (2000) Intrinsic
respectively). However, in contrast to the present data, the predator-
properties of lateral and dorsolateral columns of periaqueductal
elicited flight behaviour was facilitated by 5 mg/kg and 10 mg/kg
FLX (Griebel et al. 1996; Blanchard et al. 1997). Therefore, albeit
Blackwell B, Stefopoulos A, Enders P, Kuzma R, Adolphe A
similar, the substrates of DPAG-evoked and predator-elicited
(1978) Anticholinergic activity of two tricyclic antidepressants.
flight behaviours seem not to be identical. However, in contrast to
the lack of effects of the higher doses of CLM (>10 mg/kg) in
Blanchard RJ, Griebel G, Henrie JA, Blanchard DC (1997)
both DPAG-evoked innate and shuttle-box escape behaviours,
Differentiation of anxiolytic and panicolytic drugs by effects
CLM (15 mg/kg) attenuated the bar-pressing decremental escape
on rat and mouse defense test batteries. Neurosci Biobehav
in which rats learn to titrate the intensity of DPAG stimulation
(Kiser et al. 1978). Decremental escape was further potentiated by
Blok BF, Holstege G (1996) The neuronal control of micturition
para-chlorophenylalanine and depressed by 5-hydroxytryptophan,
and its relation to the emotional motor system. In: Holstege G,
supporting the 5-HT mediation of CLM attenuating effects (Kiser
Bandler R, Saper CB (eds) The emotional motor system.
and Lebovitz 1975; Kiser et al. 1978). Therefore, CLM differential
effects on DPAG-evoked, predator-elicited and bar-pressing
Croci T, Landi M, Bianchetti A, Manara L (1995) Drug-induced
escape behaviours suggest the involvement of multiple 5-HT
defaecation in rats: role of central 5-HT1A receptors. Br J
systems controlling these responses.
The pharmacology of defence reaction has long been hindered
Cusack B, Nelson A, Richelson E (1994). Binding of antidepressants
by the lack of an appropriate approach for measuring the intracranially
to human brain receptors: focus on newer generation compounds.
induced innate behaviours. Nevertheless, the present and recent
studies (Schenberg et al. 2000; Vargas and Schenberg 2001) showed
Deakin JFW, Graeff FG (1991) 5-HT and mechanisms of defence.
that the threshold logistic analysis can provide accurate information
about drug effects on the triggering level (I ), responsiveness (β)
Gentil V (1988) The aversive system, 5-HT and panic attacks. In:
) of DPAG-evoked defensive behaviours.
Simon P, Soubrié P, Wildlocher D (eds) Selected models
The neuronal mechanisms controlling each variable are not understood
of anxiety, depression and psychosis. Karger AG, Basel,
thus far; however, either distinct receptors or ion channels of the
same or different neuron types could underlie eventual changes in
Graeff FG (1981) Minor tranquilizers and brain defense systems.
these parameters (Schenberg et al. 2000). More importantly, the
method herein employed was sensitive to behaviour-specific drug
Gray JA (1991) The psychology of fear and stress, 2nd edn.
effects that are often masked in operant analysis of single active
Griebel G, Blanchard DC, Agnes RS, Blanchard RJ (1995)
Nashold BS, Wilson WP, Slaughter DG (1969) Sensations evoked
Differential modulation of antipredator defensive behavior in
by stimulation in the midbrain of man. J Neurosurg 30:14–24
Swiss- Webster mice following acute or chronic administration
Nogueira RL, Graeff FG (1995) Role of 5-HT receptor subtypes in
of imipramine and fluoxetine. Psychopharmacology 120:57–66
the modulation of dorsal periaqueductal gray generated
Griebel G, Blanchard DC, Blanchard RJ (1996) Predator-elicited
aversion. Pharmacol Biochem Behav 52:1–6
flight responses in Swiss-Webster mice: an experimental
Paxinos G, Watson C (1986) The rat brain in stereotaxic
model of panic attacks. Prog Neuropsychopharmacol Biol
coordinates, 2nd edn. San Diego, Academic Press
Ramos RT, Gentil V, Gorenstein C (1993) Clomipramine and
Jenck F, Broekkamp CLE, Van Delft AML (1989) Effects of
initial worsening in panic disorder: beyond the “jitteriness
serotonin receptor antagonists on PAG stimulation induced
syndrome”. J Psychopharmacol 7:265–269
aversion: different contributions of 5HT , 5HT
Richelson E, Divinetz-Romero S (1977) Blockade by psychotropic
receptors. Psychopharmacology 97:489–495
drugs of the muscarinic acetylcholine receptor in cultured
Jenck F, Broekkamp CLE, Van Delft AML (1990) The effect of
nerve cells. Biol Psychiatry 12:771–785
antidepressants on aversive periaqueductal gray stimulation in
Sanger GJ (1996) 5-Hydroxytryptamine and functional bowel
disorders. Neurogastroenterol Motil 8:319–331
Jenck F, Moreau JL, Martin JR (1995) Dorsal periaqueductal
Sanger GJ, Banner SE, Smith MI, Wardle KA (1998) SB-207266:
grayinduced aversion as a simulation of panic anxiety: elements
5-HT receptor antagonism in human isolated gut and prevention
of face and predictive validity. Psychiatry Res 57:181–191
of 5-HT-evoked sensitization of peristalsis and increased
Jenck F, Martin JR, Moreau JL (1996) Behavioral effects of
defaecation in animal models. Neurogastroenterol Motil 10:
CCKB receptor ligands in a validated simulation of panic
anxiety in rats. Eur J Neuropsychopharmacol 6:291–298
Schenberg LC, Graeff FG (1978) Role of the periaqueductal gray
Jenck F, Moreau JL, Berendsen HH, Boes M, Broekkamp CL,
substance in the antianxiety action of benzodiazepines.
Martin JR, Wichmann J, Van Delft AM (1998) Antiaversive
receptor agonists and fluoxetine in a model
Schenberg LC, Costa MB, Borges PCL, Castro MFS (1990)
of panic-like anxiety in rats. Eur Neuropsychopharmacol
Logistic analysis of the defense reaction induced by electrical
stimulation of the rat mesencephalic tectum. Neurosci Biobehav
Kiser RS, Lebovitz RM (1975) Monoaminergic mechanisms in
aversive brain stimulation. Physiol Behav 15:47–53
Schenberg LC, Marçal LPA, Seeberger F, Barros MR, Sudré ECM
Kiser RS, German DC, Lebovitz RM (1978) Serotonergic reduction
(2000) L-type calcium channels selectively control the
of dorsal central gray area stimulation-produced aversion.
defensive behaviors induced by electrical stimulation of dorsal
periaqueductal gray and overlying collicular layers. Behav
Lovick TA (1994) Influence of the dorsal and median raphe nuclei
on neurons in the periaqueductal gray matter: role of
Schütz MTB, Aguiar JC, Graeff FG (1985) Anti-aversive role of
5-hydroxytryptamine. Neuroscience 59:993–1000
serotonin in the dorsal periaqueductal grey matter. Psycho-
Luppi PH, Aston-Jones G, Akaoka H, Chouvet G, Jouvet M
(1995) Afferent projections to the rat locus coeruleus
Shiekhattar R, Aston-Jones G (1993) Sensory responsiveness of
demonstrated by retrograde and anterograde tracing with
brain noradrenergic neurons is modulated by endogenous
cholera-toxin B subunit and Phaseolus vulgaris leucoagglutinin.
Snyder SH, Yamamura HI (1977) Antidepressants and the
Lydiard RB, Falsetti SA (1999) Experience with anxiety and
muscarinic acetylcholine receptor. Arch Gen Psychiatry 34:
depression treatment studies: implications for designing irritable
bowel syndrome clinical trials. Am J Med 107:65S–73S
Sohn UD, Kim CY (1997) Suppression of the rat micturition
Lydiard RB, Laraia MT, Howell EF, Ballenger JC (1986) Can
reflex by imipramine. J Auton Pharmacol 17:35–41
panic disorder present as irritable bowel syndrome? J Clin
Sudré ECM, Barros MR, Sudré GN, Schenberg LC (1993)
Thresholds of electrically induced defence reaction of the rat:
Lydiard RB, Greenwald S, Weissman MM, Johnson J, Drossman
short- and long-term adaptation mechanisms. Behav Brain Res
DA, Ballenger JC (1994) Panic disorder and gastrointestinal
symptoms: findings from the NIMH Epidemiologic Catchment
Valentino RJ, Page ME, Luppi PH, Zhu Y, Van Bockstaele E,
Area project. Am J Psychiatry 151:64–70
Aston-Jones G (1994) Evidence for widespread afferents to
Miczek KA (1973) Effects of scopolamine, amphetamine and
Barrington’s nucleus, a brainstem region rich in corticotropin-
chlordiazepoxide on punishment. Psychopharmacologia 28:
releasing hormone neurons. Neuroscience 62:125–143
Valentino RJ, Kosboth M, Colflesh M, Miselis RR (2000) Trans-
Miyata K, Kamato T, Nishida A, Ito H, Yuki H, Yamano M,
neuronal labeling from the rat distal colon: anatomic evidence
Tsutsumi R, Katsuyama Y, Honda K (1992) Role of the serotonin
for regulation of distal colon function by a pontine corticotropin-
receptor in stress-induced defecation. J Pharmacol Exp Ther
releasing factor system. J Comp Neurol 417:399–414
Vargas LC, Schenberg LC (2001) Long-term effects of clomipramine
Miyata K, Ito H, Fukudo S (1998) Involvement of the 5-HT
and fluoxetine on dorsal periaqueductal gray-evoked innate
receptor in CRH-induced defecation in rats. Am J Physiol
defensive behaviours of the rat. Psychopharmacology 155:
Monnikes H, Schmidt BG, Tebbe J, Bauer C, Tache Y (1994)
Vargas LC, Marques TA, Schenberg LC (2000) Micturition and
Microinfusion of corticotropin releasing factor into the locus
defensive behaviors are controlled by distinct neural networks
coeruleus/subcoeruleus nuclei stimulates colonic motor function
within the dorsal periaqueductal gray and deep gray layer of
the superior colliculus of the rat. Neurosci Lett 280:45–48
1.0 Flora The flora surveyed at the site was done so using stratified sampling and random sampling. The sample points are shown in Figure 3.01. In Table 1.1 that follows, there is a complete of the plants and trees found at the site. FLORA: Table 1- A Complete List of the Floral Species Found at the Vision City Project Site Species Common Name Species Scientific Name Wild
FINGER LAKES RUNNERS CLUB Wednesday, June 13, 2007 Martha Van Rensselaer Hall, Rushmore Conference Room (114 MVR), Cornell University Meeting commenced at 5:43 pm Board members present: Casey Carlstrom, Karen Grover, Joe Reynolds, Chris Reynolds, Steve Shaum, Katie Stettler, Nancy Kleinrock, Lorrie Tily, Evan Kurtz, Alan Lockett, Jim Miner, John Dailey, Jr., Tim Ingall, Don Tily
 and post-drug threshold curves of each group were modelled bymeans of indicator variables and compared for either location(triggering level) or parallelism (responsiveness) using likelihood-ratio coincidence tests. Behaviour triggering level was representedby estimates of median intensity (I ±SE). Behavioural respon-
siveness was represented by regression curvature (β±SE), i.e. theparameter that governs the rate of change in response probabilityas a function of stimulus change. Finally, whenever a low responseoutput precluded the logistic fitting, maximum accumulatedfrequencies (P
using odds ratio (ψ) and Pearson’s χ2 analyses. A comprehensivedescription of our original approach to the threshold logistic analysisof intracranially induced behaviours can be found elsewhere(Schenberg et al. 2000).
and post-drug threshold curves of each group were modelled bymeans of indicator variables and compared for either location(triggering level) or parallelism (responsiveness) using likelihood-ratio coincidence tests. Behaviour triggering level was representedby estimates of median intensity (I ±SE). Behavioural respon-
siveness was represented by regression curvature (β±SE), i.e. theparameter that governs the rate of change in response probabilityas a function of stimulus change. Finally, whenever a low responseoutput precluded the logistic fitting, maximum accumulatedfrequencies (P
using odds ratio (ψ) and Pearson’s χ2 analyses. A comprehensivedescription of our original approach to the threshold logistic analysisof intracranially induced behaviours can be found elsewhere(Schenberg et al. 2000).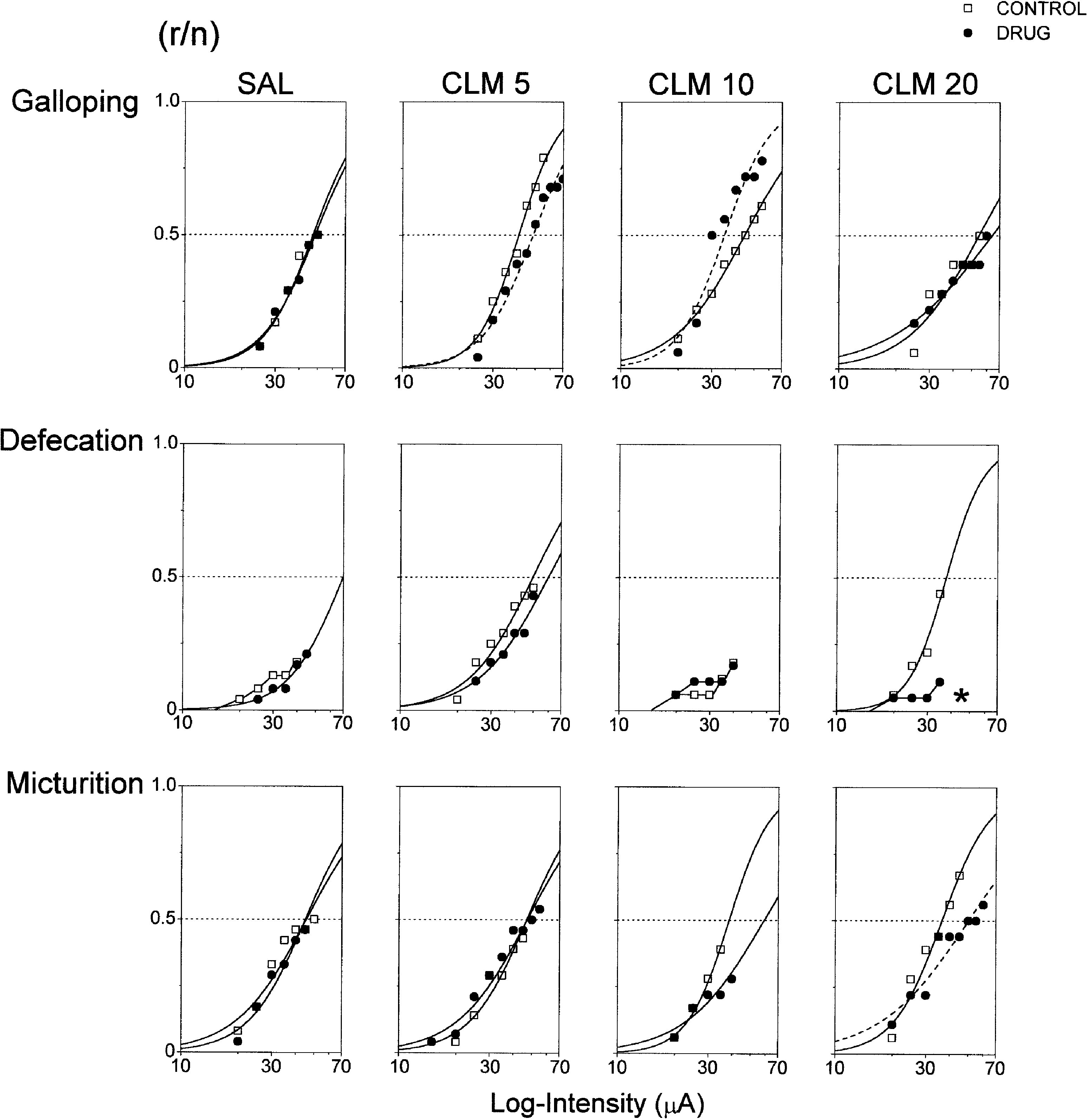 Fig. 2 Acute effects of
Fig. 2 Acute effects of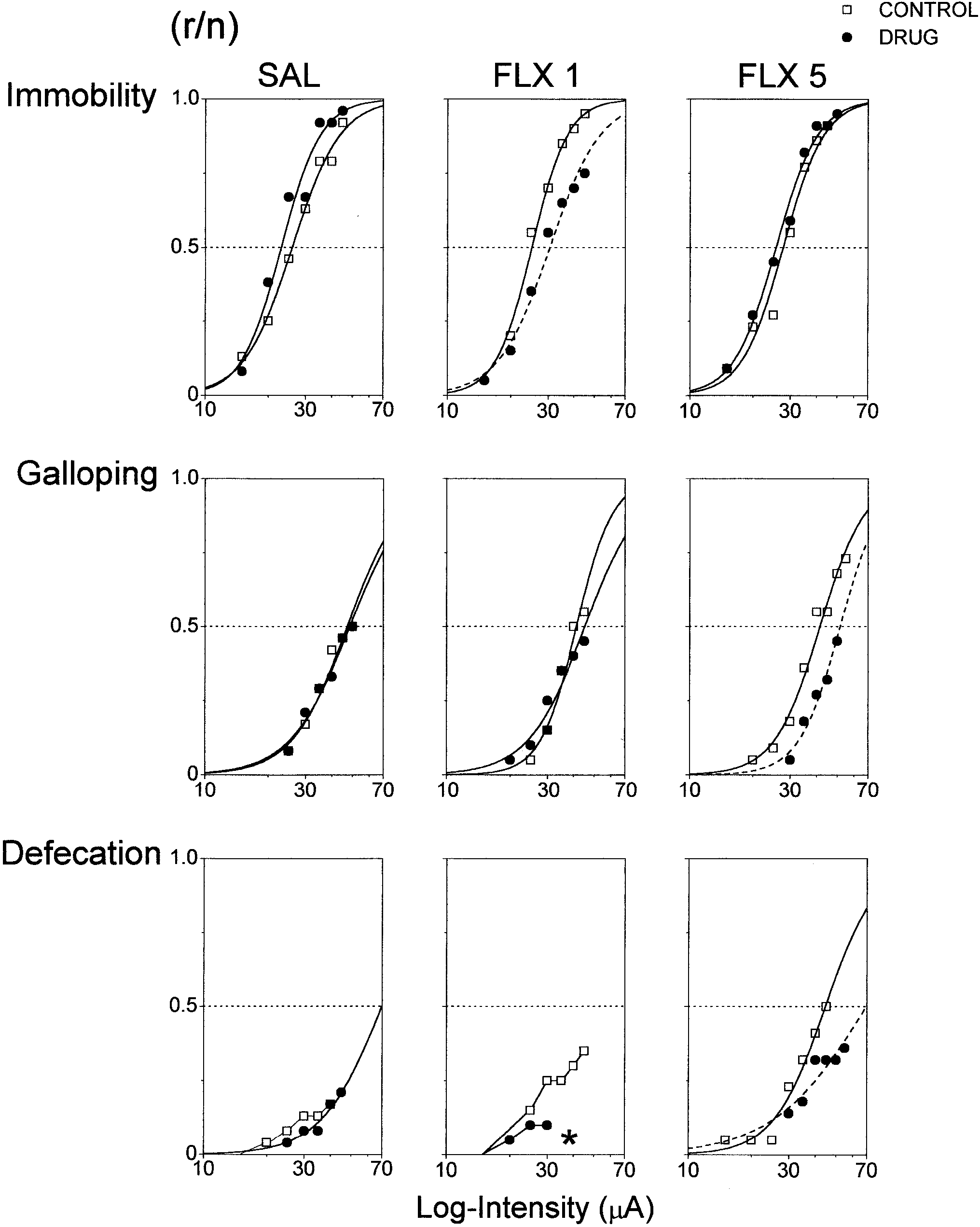
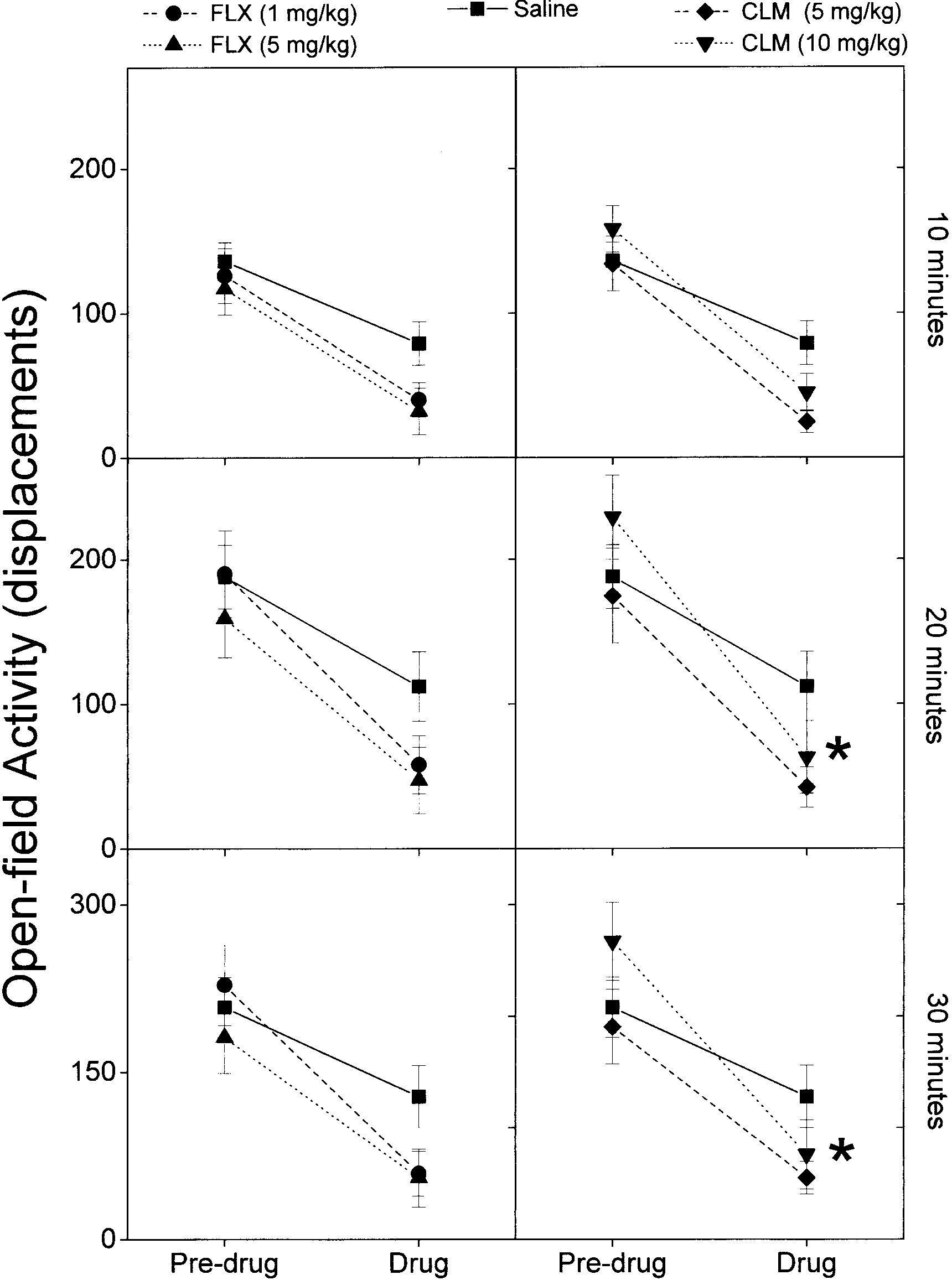 Fig. 4 Fluoxetine (FLX 1 mg/kg and 5 mg/kg, i.p.) and clomipramine
Fig. 4 Fluoxetine (FLX 1 mg/kg and 5 mg/kg, i.p.) and clomipramine