Kamagra enthält Sildenafilcitrat als pharmakologisch aktiven Bestandteil. Dieser hemmt selektiv die Phosphodiesterase-5 und erhöht dadurch die Konzentration von cGMP im Corpus cavernosum. Der Effekt ist zeitlich begrenzt, da die Halbwertszeit von Sildenafil etwa vier Stunden beträgt. In der galenischen Form als Mundgel erfolgt die Resorption besonders rasch, was zu einem schnelleren Wirkeintritt führt. Der Abbau erfolgt überwiegend hepatisch über CYP3A4, wobei ein aktiver Metabolit entsteht, der zur Gesamtwirkung beiträgt. Typische Nebenwirkungen ergeben sich aus der Vasodilatation, darunter leichte Kopfschmerzen und nasale Kongestion. In klinischen Beschreibungen wird kamagra oral jelly im Zusammenhang mit der schnelleren Absorption erwähnt.
Inactivation of hippocampus or caudate nucleus with lidocaine differentially affects expression of place and response learning
NEUROBIOLOGY OF LEARNING AND MEMORY 65, 65 – 72 (1996) Article No. 0007
Inactivation of Hippocampus or Caudate Nucleus with Lidocaine
Differentially Affects Expression of Place and Response Learning
MARK G. PACKARD* AND JAMES L. MCGAUGH†,1
*Department of Psychology, University of New Orleans 70148; and †Center for the Neurobiology of Learning and Memoryand Department of Psychobiology, University of California, Irvine 92717
response learning, respectively (2), in a visually cued ex-
Involvement of the hippocampus and caudate nucleus in
place and response learning was examined by functionally
learning is acquired faster than caudate-dependent re-
inactivating these brain regions bilaterally with infusions
sponse learning, and (3) when animals shift to caudate-
of lidocaine. Rats were trained to approach a consistently
dependent response learning with extended training, the
baited arm in a cross-maze from the same start box (four
hippocampal-based place representation remains intact.
trials/day/14 total days). On Days 8 and 16 a single probe
trial was given, in which rats were placed in the start boxopposite that used in training and allowed to approach amaze arm. Three minutes prior to the probe trial, rats
received bilateral injections of either saline or a 2% lido-caine solution (in order to produce neural inactivation)
Throughout the history of research on animal
into either the dorsal hippocampus or dorsolateral caudate
learning there have been conflicting views concern-
nucleus. On the probe trials, rats which entered the baited
ing the fundamental issue of what animals learn.
maze arm (i.e., approached the place where food was lo-
Cognitive theorists such as Tolman (1932; 1948) pro-
cated during training) were designated place learners, and
posed that animals acquire knowledge of ‘‘what-
rats which entered the unbaited maze arm (i.e., made the
leads-to-what’’ that result in expectations of the con-
same turning response as during training) were desig-
sequences of their behavior. In contrast, following
nated response learners. Saline-treated rats displayed
the pioneering work of Thorndike (1898; 1933) and
place learning on the Day 8 probe trial and response learn-
Pavlov (1927), other theorists proposed that animal
ing on the Day 16 probe trial, indicating that with ex-
learning consists of the formation of stimulus – re-
tended training there is a shift in learning mechanisms
sponse (S-R) habits (Hull, 1943; Guthrie, 1935) or,
controlling behavior. Rats given lidocaine injections intothe hippocampus showed no preference for place or re-
simply, the learning of motor responses.
sponse learning on the Day 8 probe trial, but displayed
In experiments addressing this issue, rats were
response learning on the Day 16 probe trial, indicating a
trained in mazes and subsequently tested in a vari-
blockade of place learning following inactivation of the
ety of ways in an attempt to discover whether the
hippocampus. Rats given lidocaine injections into the cau-
learning was based on acquisition of knowledge or
date nucleus displayed place learning on both the Day 8
learning of responses (e.g., Tolman, Ritchie, & Kal-
and the Day 16 probe trials, indicating a blockade of re-
ish, 1946, 1947; Blodgett & McCutchan, 1947, 1948;
sponse learning following inactivation of the caudate nu-
Blodgett, McCutchan, & Matthews, 1949; Thomp-
cleus. The findings indicate: (1) the hippocampus and cau-
son & Thompson, 1949; Hill & Thune, 1952; Schar-
date nucleus selectively mediate expression of place and
lock, 1955). For example, after animals were trainedto enter one arm of a T-maze where they were re-
1 This research was supported by NRSA Grant 1 F32NS08973-
warded with food in a goal box, had they learned a
01 (M.G.P.) and USPHS Grant MH12526 from NIDA and NIMH
turning response or, alternatively, had they learned
(J.L.M.). The authors thank Norman White, Norman Wienberger,
where food was located? In order to examine this
and Richard Thompson for helpful comments on an earlier version
question, Tolman and colleagues introduced the
of the manuscript. Address correspondence and reprint requests
cross-maze paradigm (Tolman et al., 1946, 1947).
to Mark G. Packard, Department of Psychology, University ofNew Orleans, New Orleans, LA 70148.
The cross-maze is essentially a T-maze built such
Copyright ᭧ 1996 by Academic Press, Inc.
All rights of reproduction in any form reserved.
that the choice point connecting the two goal boxes
Findings of double dissociations of the effects of
(e.g., east – west), can be approached from start boxes
brain lesions and intracerebral drug treatments on
located on either side of the maze (e.g., north –
memory strongly support the hypothesis that the
south). During an initial training period, animals
hippocampal system and caudate nucleus mediate
are placed in the same start box and trained on sev-
different forms of memory. Lesions of the hippocam-
eral trials to obtain a food pellet from the same goal
pal system (Packard, Hirsh, & White, 1989; Pack-
box. Following this training period, a critical probe
ard & McGaugh, 1992; McDonald & White, 1993;
trial is given in which animals are placed in the
Kesner, Bolland, & Dakis, 1993), and posttraining
start box opposite the one used during training and
injections of drugs into the hippocampus (Packard &
allowed to approach either goal box. According to
White, 1991; Packard, Cahill, & McGaugh, 1994),
cognitive theory, animals should approach the goal
alter cognitive learning but do not affect response
box which was baited during training, since they
learning. Conversely, lesions of the caudate nucleus
acquired information concerning the spatial location
(Packard et al., 1989; Packard & McGaugh, 1992;
of the food reward. Animals displaying this behavior
McDonald & White, 1993; Kesner et al., 1993), and
on the probe trial are designated ‘‘place learners.’’
posttraining injections of drugs into the caudate
In contrast, S-R theory predicts that animals should
(Packard & White, 1991; Packard et al., 1994),
approach the goal box which was unbaited during
alter response learning but do not affect cognitive
training, since they have learned a response ten-
dency (i.e., a specific body turn at the choice point).
The present study was designed to determine
Animals displaying this behavior are designated ‘‘re-
whether the evidence suggesting differential roles of
sponse learners.’’ Although the cross-maze paradigm
the hippocampus and caudate nucleus in memory
provides a procedure to contrast cognitive and S-R
might provide a resolution of the place versus re-
learning theories, the findings of many experiments
sponse learning question. Rats implanted bilaterally
using this task were inconclusive. Under some condi-
with cannulae in either the hippocampus or caudate
tions animals indicated knowledge of the location of
nucleus were first trained (four trials per day) using
rewards (i.e., place learning), and under other condi-
one starting alley of a cross-maze (e.g., south) and
tions they made specific body turn responses on the
consistently rewarded in the goal box of one alley
probe trial (for review see Restle, 1957).
(e.g., west). On Days 8 and 16 the rats were given a
Findings indicating that both place and response
single test trial using the other starting alley (e.g.,
learning occurred raises the possibility that these two
north) to determine whether they made the same
forms of learning might be mediated by distinct neu-
turning response made in training or made a differ-
ral mechanisms. This hypothesis is consistent with
ent turning response and went to the place where
extensive evidence suggesting that different forms of
food was located during the training. Prior to the
learning and memory are mediated by different neu-
test trials lidocaine or saline was infused bilaterally
ral systems in humans (e.g., Milner, 1962; Corkin,
into either the caudate nucleus or hippocampus. Li-
1965; Weiskrantz & Warrington, 1979; Cohen &
docaine produces a temporary functional blockade of
Squire, 1980; Schacter, 1992), monkeys (e.g., Gaffan,
neural activity following local administration in the
1974; Zola-Morgan, Squire, & Mishkin, 1982; Ma-
brain and has been used in studies examining brain
hut & Moss, 1984), and rats (e.g., Hirsh, 1974;
function and memory (e.g., Perez-Ruiz & Prado-Al-
O’Keefe & Nadel, 1978; Olton, Becker, & Handel-
cala, 1989; Salinas, Packard, & McGaugh, 1993). If
mann, 1979; Sutherland & Rudy, 1989). Within the
cognitive and response learning occur in parallel and
context of the debate between cognitive and S-R
are mediated by different neural systems, selective
learning theorists, it has been suggested that the hip-
temporary inactivation of one of these neural sys-
pocampal system may selectively mediate ‘‘cognitive’’
tems prior to testing should influence the type of
memory (Hirsh, 1974; Mishkin & Petri, 1984), and
learning, i.e., place or response learning expressed
that the caudate nucleus may selectively mediate ‘‘S-
R habit’’ formation (Mishkin & Petri, 1984; Packardand White, 1987; Packard, Hirsh, & White, 1989).
Similarly, Kesner and colleagues hypothesized thatthe hippocampus selectively mediates allocentric spa-
tial behavior within a ‘‘data-based’’ memory system,while the caudate nucleus selectively mediates ego-
The subjects were 50 male Sprague – Dawley rats
centric learning within an ‘‘expectancy-based’’ mem-
(275 – 300 g). Animals were individually housed in a
ory system (Kesner & DiMattia, 1987).
temperature-controlled environment on a 12-h light/
INACTIVATION OF HIPPOCAMPUS OR CAUDATE NUCLEUS
dark cycle with the lights on from 7 AM to 7 PM. All
intracerebral administration (Parent & McGaugh,
animals were given ad lib. access to water.
1993). The injections were delivered over a period of37 s using a syringe pump (Sage Instruments), and
the injection needles (extending 1 mm from the endof the guide cannulae) were left in place an addi-
The apparatus was a wooden cross maze painted
tional 60 s to allow for diffusion of the solution away
flat gray. The maze consisted of four arms (north,
from the needle tip. Lidocaine injections were ad-
south, east, and west) of an eight-arm radial maze
ministered approximately 2 – 3 min prior to the
(other maze arms were removed). The arms of the
probe trial tests on Days 8 and 16 of training. Saline
cross maze measured 60 1 9 cm. The center platform
injections were administered using procedures iden-
of the maze connecting the four arms measured 40
tical to those used for lidocaine injections.
cm in diameter. A clear Plexiglas cross-shaped alley-way structure placed on the center platform of the
modified radial maze connected the four arms of thecross-maze. The alleyways measured 20 1 9 1 15
At the completion of behavioral training, animals
cm. A recessed food well was present at the end of
were deeply anesthetized with a 1.0-ml injection of
the west arm of the maze. The maze was located in
sodium pentobarbital and perfused with saline fol-
a testing room that contained many extramaze cues
lowed by 10% formal-saline. The brains were re-
including wall posters, a lamp, table, animal cage
moved and subsequently sectioned at 20-mm sections
through the cannula tract region and stained withCresyl violet. Cannula placements were examined
for verification of needle tip location using the atlasof Paxinos and Watson (1986).
Animals were anesthetized with sodium pentobar-
Cannulae placements for both dorsolateral cau-
bital (50 mg/kg) and implanted with bilateral guide
date nucleus and dorsal hippocampus are shown in
cannula in the dorsal hippocampus (10 mm length)
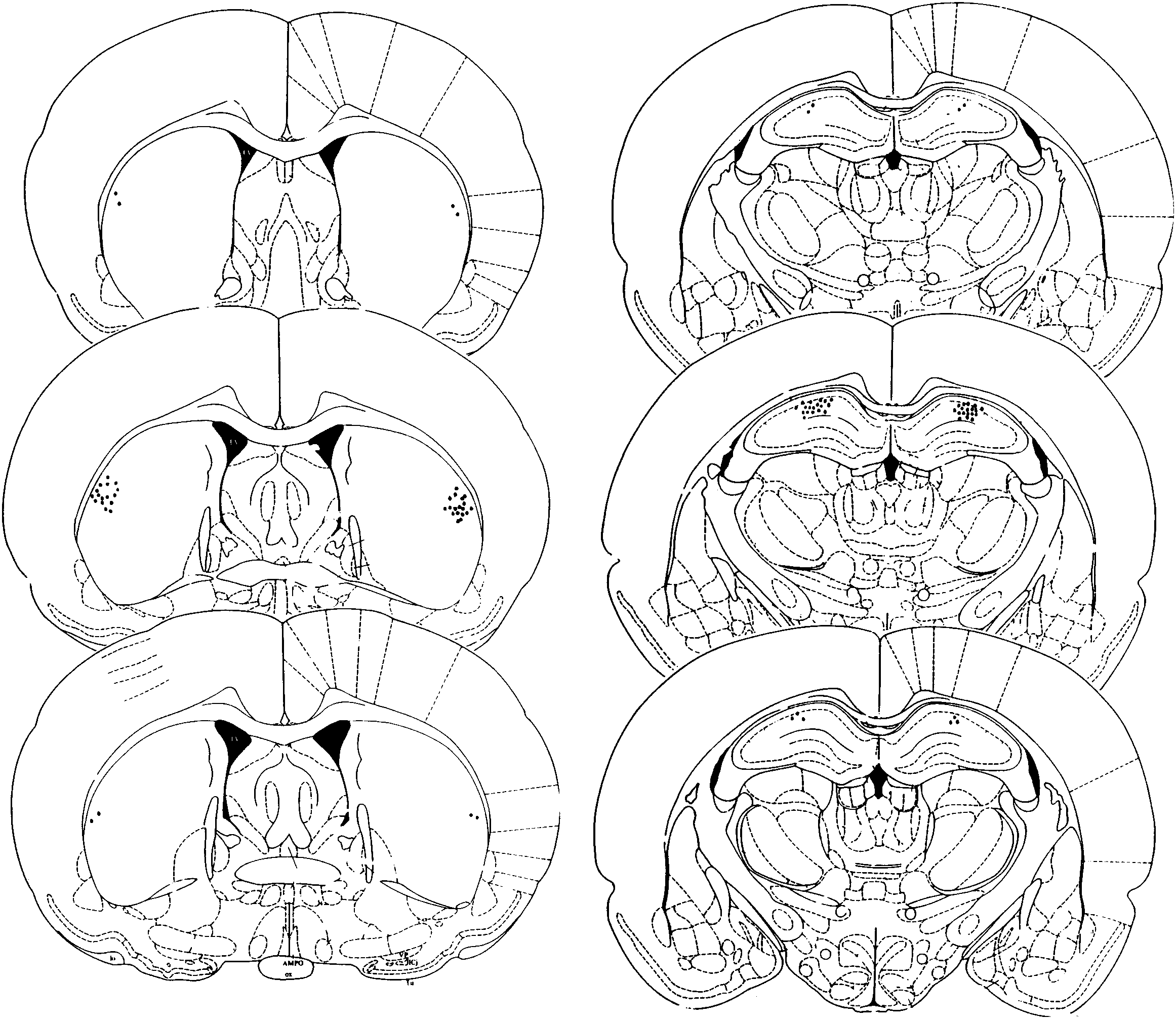
Fig. 1 (left, right, respectively). Caudate nucleus
or dorsolateral caudate nucleus (15 mm length) us-
placements were located in the dorsolateral caudate,
ing standard stereotaxic techniques. The cannulae
ranging from 0.20 mm to 00.30 mm from bregma
(23 gauge) were anchored to the skull with jewelers
(Fig. 1, left). Hippocampal placements were located
screws and dental acrylic. Coordinates for the dorsal
in the dorsal hippocampus, ranging from 02.8 mm
hippocampal placements were AP Å 03.1 mm, ML
to 03.3 mm from bregma (Fig. 1, right).
Å {2.5 mm, and DV Å 02.0 mm from bregma. Coor-dinates for the dorsolateral caudate nucleus place-
ments were AP Å 0.26 mm, ML Å {4.2 mm, DVÅ 04.0 mm. After surgery, stylets (30 gauge) were
Prior to training, all rats were reduced to 85% of
inserted and left in place to ensure cannulae patency
ad lib. body weights over 7 days and maintained at
until injections were made. Behavioral testing began
this weight throughout the experiment. On 2 consec-
utive days, rats were placed into the cross-maze inthe start box (south arm) and allowed to explore the
maze for 5 min. No food was present in the maze on
A 2% lidocaine hydrochloride solution (Western
either of these 2 habituation days. Access to the
Medical Supply, Inc.) was used to produce reversible
north arm of the cross-maze was blocked during ha-
inactivation of brain sites. Injections (0.5 ml) were
bituation sessions and the subsequent food rewarded
administered intracerebrally using 30-gauge injec-
training trials with a clear Plexiglas shield. Follow-
tion needles inserted into the guide cannulae. The
ing habituation on both days, the animals were al-
needles were connected by polyethylene tubing to
lowed to consume ten 45-mg Noyes food pellets in
10-ml Hamilton microsyringes (Hamilton Co., Reno,
their home cage. Food trials began on Day 3. On
NV). The volume of 2% lidocaine solution used was
each food trial rats were placed into the start box
chosen on the basis of previous evidence indicating
and allowed to traverse the maze and consume a
that this volume produces functional inactivation of
single Noyes food pellet located in the food cup at
the caudate nucleus (Perez-Ruiz & Prado-Alcala,
the end of the goal arm of the cross maze (west arm).
1989) and amygdala (Salinas et al., 1993) sufficient
On the initial food trial only, a trail of four pellets
to cause memory impairment. Other findings indi-
leading to the food cup was placed along the length
cate that volumes of 2% lidocaine solution as low as
of the goal arm. Each rat received four food rewarded
0.25 ml can produce memory impairment following
trials per day. Entries into the unbaited arm of the
Dorsolateral caudate nucleus (left) and dorsal hippocampus (right) cannulae placements showing the anterior/posterior extent
of needle tip locations at 0.5-mm sections. Caudate nucleus placements ranged from 0.20 to 0.30 mm from bregma. Hippocampalplacements ranged from 02.8 to 03.0 mm from bregma. (Plates adapted from atlas of Paxionos and Watson, 1986).
cross-maze (east) were scored as incorrect responses
On Day 8 of the food-rewarded trials, rats were
during the training trials, and entries into the baited
assigned to experimental groups (n Å 12–14 per
arm of the cross-maze (west) were scored as correct
group) in a rank-order method to assure that levels
responses. A correction procedure was used such
of learned performance (i.e., number of correct re-
that rats making an incorrect response were allowed
sponses over the first 7 days of training) of the groups
to trace back to the baited maze arm and consume
were comparable prior to treatment. On Day 8 a sin-
the food pellet. If a rat failed to consume the food
gle probe trial was given. Three minutes prior to the
pellet within 2 min, the trial was terminated and
probe trial, half of the animals with caudate nucleus
the rat was manually placed in the goal box and
implants received a saline injection and the other half
allowed to consume the pellet. After consuming the
received a lidocaine injection. Similarly, half of the
pellet on a given trial, the rat was placed in a holding
animals with hippocampal implants received a saline
cage located directly behind the start arm for a 30-
injection prior to the probe trial, and half received an
injection of lidocaine. On the probe trial, animals
INACTIVATION OF HIPPOCAMPUS OR CAUDATE NUCLEUS
the Day 8 probe trial, animals given saline injectionsinto either the hippocampus or caudate nucleus werepredominantly place learners (hippocampus – salinex2 Å 7.14, p õ .05; caudate nucleus – saline x2 Å5.34, p õ .05). Rats given intracaudate injections oflidocaine were also predominantly place learners (x2Å 5.34, p õ .05), while those receiving intrahippo-campal injections of lidocaine did not show a signifi-cant trend toward either place or response learning(x2 Å 0, n.s.). These findings indicate that functionalinactivation of the hippocampus, but not caudate nu-cleus, prevented the expression of place learning.
On the Day-16 probe trial, animals given saline
injections into either the hippocampus or caudatenucleus were predominantly response learners (hip-pocampus – saline x2 Å 4.58, p õ .05; caudate nu-cleus – saline x2 Å 5.34, p õ .05). Rats receiving in-trahippocampal injections of lidocaine were also pre-dominantly response learners (x2 Å 5.34, p õ .05),
Number of rats in each treatment group that exhib-
while those receiving intracaudate injections of lido-
ited place or response learning on both the Day 8 and Day 16 test
caine were predominantly place learners (x2 Å 8.32,
p õ .05). These findings indicate that with extendedtraining, saline-treated rats switched from dis-playing place learning to response learning. Func-
were placed into the start box opposite that used dur-
tional inactivation of the caudate nucleus, but not
ing training (i.e., north arm) and were allowed to
hippocampus, prevented the expression of response
make a single entry into either the baited or the un-
learning and, in fact, preserved place learning.
baited maze arm. The entrance to the south maze arm(i.e., the arm containing the start box used during
training) was blocked by a clear Plexiglas shield onthe probe trials. Animals entering the baited arm on
The results indicate that place and response learn-
the probe trial were designated ‘‘place’’ learners (i.e.,
ing occur concurrently and are mediated by different
animals going to the place where food was located
neural systems involving the hippocampus and cau-
during training), and animals entering the unbaited
date nucleus, respectively. The findings are consis-
arm on the probe trial were designated ‘‘response’’
tent with previous studies using pretraining irre-
learners (i.e., animals making the same turning re-
versible lesions of the caudate nucleus (Thompson,
Guilford, & Hicks, 1980), and fimbria – fornix (Decas-
On Day 9 of training, food-rewarded training (four
tro, 1974), which reported that such lesions result in
trials per day) was reinstated using procedures iden-
a predominant tendency toward place and response
tical to those of Training Days 1 – 7. On Day 16, a
learning, respectively. Our findings indicate that in
second probe trial was given using procedures identi-
addition to acquisition, the functional integrity of
cal to the probe trial given on Day 8. Animals re-
the hippocampus and caudate nucleus are necessary
ceived the same intracerebral saline or lidocaine
for the expression of these two forms of learned be-
treatments given on the Day 8 probe trial and were
again allowed to make a single entry into either the
The finding that rats initially acquire place infor-
mation and that the expression of such learning isblocked by inactivation of the hippocampus is consis-
tent with extensive evidence implicating the hippo-campus in spatial learning (e.g., O’Keefe & Nadel,
Figure 2 shows the results of the test trials on
Days 8 and 16. x2 analyses (p õ .05 for all compari-
The hippocampal injection sites in the present
sons) were computed in order to determine whether
study were located along the dorsal edge of the hip-
groups showed a significant tendency to display
pocampus, and areas of parietal cortex located dorso-
place or response learning on the probe trials. On
lateral to the lidocaine injection sites may also be
involved in acquisition of spatial information. For
rats, a response learning tendency had apparently
example, large lesions of parietal cortex selectively
not been acquired by this system prior to the eighth
impair acquisition of a spatial cheese-board task
day of training. Thus, one difference between the
(Kesner, Farnsworth, & DiMattia, 1989) and pro-
operating characteristics of the hippocampal-based
duce deficits in acquisition and retention of spatial
and caudate-based memory systems is that the for-
behavior in the Morris water maze (DiMattia &
mer system appears to be involved in the rapid ac-
Kesner, 1988). However, the posterior extent of the
quisition of new information, while the latter
large parietal cortex lesions reported to impair spa-
system acquires information in a slower, incremen-
tial behavior (Kesner, Farnsworth, & DiMattia,
1989) is approximately 0.5 – 0.8 mm anterior and
With extended training in the cross-maze, animals
1.0 – 1.2 mm lateral to our effective injection sites in
acquired a response learning tendency (i.e., on the
dorsal hippocampus. Furthermore, small lesions of
Day 16 probe trial control rats made the same turn-
parietal cortex which may more closely mimic the
ing response as during training). The finding that
behavioral effects of any limited spread of lidocaine
the animals shifted from place learning to response
from hippocampus in the present study do not im-
learning with increased training is consistent with
pair acquisition of spatial behavior in the Morris
that of previous research (e.g., Ritchie, Aeschli-
water maze or the radial maze (Kolb, Sutherland, &
man, & Pierce, 1950; Hicks, 1964). Importantly, se-
Whishaw, 1983). Thus, although the possibility that
lective functional inactivation of the caudate nucleus
the spread of lidocaine to adjacent parietal cortex
prevented the expression of the acquired response
influenced the expression of place learning cannot
and revealed preserved place learning. Thus, the pre-
be completely ruled out, the effect of lidocaine injec-
dominance of response learning expressed after ex-
tions on place learning observed in the present study
tensive training is not due to elimination of the place
was likely due to neural inactivation of the hippo-
representation. These findings provide strong evi-
dence that the two memory systems are functioning
The finding that the expression of response learn-
ing is blocked by functional inactivation of the cau-
In 1957, Restle proposed a resolution of the place
date nucleus is consistent with the view that the
versus response learning debate based on findings
caudate nucleus selectively mediates learning in-
suggesting that either type of learning could be pre-
volving the use of ‘‘egocentric’’ cues to guide naviga-
dicted based on the availability of various ex-
tion (Potegal, 1972; Kesner & DiMattia, 1987). For
tramaze/intramaze cues present (Restle, 1957).
example, lesions of the caudate nucleus impair ac-
Thus, in open mazes located in visually heteroge-
quisition of left – right maze discrimination tasks
neous environments place learning is dominant
(Cook and Kesner, 1988; Kesner et al., 1993). As
(Blodgett & McCutchan, 1947; Blodgett, McCut-
the region of the caudate targeted by the lidocaine
chan, & Matthews, 1949; Tolman et al., 1946, 1947),
injections (dorsolateral) receives cortical input from
while in closed mazes or visually homogeneous envi-
the somatosensory cortex (Heimer, Alheid, & Zar-
ronments response learning is dominant (Blodgett &
bosky, 1985) such input may be critical for establish-
McCutchan, 1948; Thompson & Tompson, 1949;
ing an association between the maze cues and the
Hill & Thune, 1952; Scharlock, 1955). According to
egocentric cues mediating the specific body turn re-
Restle’s multiple-cue theory, a single learning mech-
quired for response learning. However, it is im-
anism was responsible for both place and response
portant to note that egocentric learning may be but
learning, and this mechanism was ‘‘tuned’’ to pro-
one example of a class of stimulus-response learning
duce either type of learning by the nature of the
functions mediated by the caudate (Packard et al.,
extramaze/intramaze environment. However, al-
1989; Packard and McGaugh, 1992; Viaud & White,
though evidence indicates that the nature of the test-
1989), each of which is hypothesized (White, 1989)
ing environment contributes to the relative expres-
to be organized on the basis of the sensory cortical
sion of place and response learning, Restle’s resolu-
input the caudate receives (Heimer et al., 1985).
tion of the debate is, at least, incomplete. The nature
Rats given saline injections into the hippocampus
of the environment appears to interact with separate
or caudate nucleus displayed a strong place learning
learning systems in determining the expression of
tendency on the Day 8 probe trial. However, animals
place or response learning. Consistent with this sug-
with the hippocampus inactivated with lidocaine
gestion, acquisition of a caudate nucleus-mediated
prior to the Day 8 probe trial did not display either
win – stay radial maze task by intact rats is facili-
place or response learning tendencies. Although the
tated when extramaze cues are reduced by sur-
caudate nucleus was presumably functional in these
rounding the maze with curtains (Packard & White,
INACTIVATION OF HIPPOCAMPUS OR CAUDATE NUCLEUS
Hirsh, R. (1974). The hippocampus and contextual retrevial of
1987). Future studies are necessary to determine
information from memory: A theory. Behavioral Biology, 12,
what other factors in addition to the nature of the
testing environment contribute to the use of a given
Hull, C. L. (1943). Principles of Behavior. Appleton-Century-
Finally, these results indicate that the place ver-
Kesner, R. P., Farnsworth, G., and DiMattia, B. D. (1989). Double
sus response controversy was based on a false prem-
dissociation of egocentric and allocentric space following me-
ise: The debate was primarily based on the assump-
dial prefrontal and parietal cortex lesions in the rat. Behav-
tion that there is only one kind of learning and that
ioral Neuroscience, 103, 956 – 961.
the appropriate test would reveal the nature of that
Kesner, R. P., Bolland, B. L., & Dakis, M. (1993). Memory for
spatial locations, motor responses, and objects: Triple dissoci-
learning (but see Tolman, 1949). Evidence that the
ation among the hippocampus, caudate nucleus, and extra-
brain uses different neural systems for acquiring
striate visual cortex. Experimental Brain Research, 93, 462 –
and expressing different kinds of learning would ap-
pear to resolve this long-lasting controversy.
Kesner, R. P., & DiMattia, B. V. (1987). Neurobiology of an attri-
bute model of memory. Progress in Psychobiology and Physio- logical Psychology, 12, 207 – 277.
Kolb, B., Sutherland, R. J., and Whishaw, I. Q. (1983). A compari-
Blodgett, H. C., & McCutchan, K. (1947). Place versus response
son of the contributions of the frontal and parietal association
learning in the simple T-maze. Journal of Experimental Psy-
cortex to spatial localization in rats. Behavioral Neuroscience,chology, 37, 412 – 422. 97, 13 – 27.
Blodgett, H. C., and McCutchan, K. (1948). The relative strength
Mahut, H., & Moss, M. (1984). Consolidation of memory: The
of place and response learning in the T-maze. Journal of
hippocampus revisited. In N. Butters and L. R. Squire (Eds.),
Comparative and Physiological Psychology, 41, 17 – 24. Neuropsychology of Memory (pp. 297 – 315). New York: Guil-
Blodgett, H. C., McCutchan, K., & Matthews, R. (1949). Spatial
learning in the T-maze: The influence of direction, turn, and
McDonald, R. J., & White, N. M. (1993). A triple dissociation of
food location. Journal of Experimental Psychology, 39, 800 –
memory systems: Hippocampus, amygdala, and dorsal stria-
tum. Behavioral Neuroscience, 107, 3 – 22.
Cohen, N. J., & Squire, L. R. (1980). Preserved learning and reten-
Milner, B. (1962). Les troubles de la memoire accompagnant des
tion of pattern analyzing skill in amnesics: Dissociation of
lesions hippocampiques bilaterales. In P. Passant (Ed.), Psys-
knowing how and knowing that. Science, 210, 207 – 210. iologie de l’Hippocampae (pp. 257 – 272). Paris: Centre de la
Cook, D., & Kesner, R. P. (1988). Caudate nucleus and memory
for egocentric localization. Behavioral and Neural Biology,
Mishkin, M., & Petri, H. L. (1984). Memories and habits: Some
49, 332 – 343.
implications for the analysis of learning and retention. In
Corkin, S. (1965). Tactually-guided maze learning in man: Effect
L. R. Squire and N. Butters (Eds.), Neuropsychology of Mem-
of unilateral cortical excision and bilateral hippocampal le-
ory (pp. 287 – 296). New York: Guilford.
sions. Neuropsychologia, 3, 339 – 351.
Mishkin, M., Malamut, B., & Bachevalier, J. (1984). Memories
DeCastro, J. M. (1974). A selective spatial discrimination deficit
and habits: Two neural systems. In G. Lynch, J. L. McGaugh,
after fornicotomy in the rat. Behavioral Biology, 12, 373 –
and N. M. Weinberger (Eds.), Neurobiology of Learning andMemory (pp. 65 – 77). New York: Guilford.
DiMattia, B. D., and Kesner, R. P. (1988). Spatial cognitive maps:
O’Keefe, J., & Nadel, L. (1978). The Hippocampus as a Cognitive
Differential roles of parietal cortex and hippocampal forma-
Map. New York: Oxford University Press.
tion. Behavioral Neuroscience, 102, 471 – 480.
Olton, D. S., Becker, J. T., & Handelmann, G. E. (1979). Hippo-
Gaffan, D. (1974). Recognition impaired and association intact
campus, space, and memory. Behavioral and Brain Sciences,
in the memory of monkeys after transsection of the fornix. 2, 313 – 365. Journal of Comparative and Physiological Psychology, 86,
Packard, M. G., & McGaugh, J. L. (1992). Double dissociation of
fornix and caudate nucleus lesions on acquisition of two water
Galanter, E. H., & Shaw, W. A. (1954). ‘‘Cue’’ vs. ‘‘reactive inhibi-
maze tasks: Further evidence for multiple memory systems.
tion’’ in place and response learning. Journal of ComparativeBehavioral Neuroscience, 106, 439 – 446. and Physiological Psychology, 47, 395 – 398.
Packard, M. G., & White, N. M. (1987). Differential roles of the
Guthrie, E. R. (1935). The Psychology of Learning. Harper and
hippocampus and caudate nucleus in memory: Selective me-
diation of ‘‘cognitive’’ and ‘‘associative’’ learning. Society for
Heimer, L., Alheid, G. F., & Zaborsky, L. (1985). Basial Ganglia. Neuroscience Abstracts, 13, 1005. In G. Paxinos, (Ed.) The Rat Nervous System (pp. 37 – 87).
Packard, M. G., & White, N. M. (1990). Lesions of the caudate
nucleus selectively impair acquisition of ‘‘reference memory’’
Hicks, L. H. (1964). Effects of overtraining on acquisition and
in the radial maze. Behavioral and Neural Biology, 53, 39 –
reversal of place and response learning. Psychological Re-ports, 15, 459 – 462.
Packard, M. G., & White, N. M. (1991). Dissociation of hippocam-
Hill, C. W., & Thune, L. E. (1952). Place and response learning
pus and caudate nucleus memory systems by posttraining
in the white rat under simplified and mutually isolated condi-
intracerebral injection of dopamine agonists. Behavioral Neu-
tions. Journal of Experimental Psychology, 43, 289 – 297. roscience, 105, 295 – 306.
Packard, M. G., Cahill, L., & McGaugh, J. L. (1994). Amygdala
theory: The role of the hippocampal formation in learning,
modulation of hippocampal-dependent and caudate nucleus-
memory, and amnesia. Psychobiology, 17, 129 – 144.
dependent memory processes. Proceedings of the National
Thompson, M. E., & Thompson, J. P. (1949). Reactive inhibition
Academy of Sciences USA 91, 8477 – 8481.
as a factor in maze learning: II. The role of reactive inhibition
Packard, M. G., Hirsh, R., & White, N. M. (1989). Differential
in studies of place learning versus response learning. Journal
effects of fornix and caudate nucleus lesions on two radial
of Experimental Psychology, 39, 883 – 891.
maze tasks: Evidence for multiple memory systems. Journal
Thompson, W. G., Guilford, M. O., & Hicks, L. H. (1980). Effects
of Neuroscience, 9, 1465 – 1472.
of caudate and cortical lesions on place and response learning in rats. Physiological Psychology, 8, 473 – 479.
Parent, M. B., & McGaugh, J. L. (1993). Time-dependent retro-
grade impairment produced by lidocaine injections into the
Thorndike, E. L. (1898). Animal Intelligence: An experimental
basolateral complex of the amygdala. Society for Neuroscience
study of the associative processes in animals. PsychologicalReview, 8, 28 – 31.
Thorndike, E. L. (1933). A proof of the law of effect. Science, 77,
Pavlov, I. P. (1927). Conditioned Reflexes. Clarendon Press, Lon-
Tolman, E. C. (1932). Purposive Behavior in Animals and Men.
Paxinos, G., & Watson, C. (1986). The Rat Brain in StereotaxicCoordinates. New York: Academic Press.
Tolman, E. C. (1948). Cognitive maps in rats and men. Psychologi-
Perez-Ruiz, C., & Prado-Alcala, R. A. (1989). Retrograde amnesia
cal Review, 56, 144 – 155.
induced by lidocaine injection into the striatum: protective
Tolman, E. C. (1949). There is more than one kind of learning.
effect of the negative reinforcer. Brain Research Bulletin, 22, Psychological Review, 55, 189 – 208.
Tolman, E. C., & Gleitman, H. (1949). Studies in spatial learning:
Potegal, M. (1972). The caudate nucleus egocentric localization
VII. Place and response learning under different degrees of
system. Acta Neurobiologica Experimentia, 32, 379 – 494.
motivation. Journal of Experimental Psychology, 39, 633 –
Restle, F. (1957). Discrimination of cues in mazes: A resolution
of the place vs. response controversy. Psychological Review,
Tolman, E. C., Ritchie, B. F., & Kalish, D. (1946). Studies in
64, 217 – 228.
spatial learning: II. Place learning versus response learning.
Ritchie, B. F., Aeschliman, B., & Pierce, P. (1950). Studies in
Journal of Experimental Psychology, 36, 221 – 229.
spatial learning: VIII. Place performance and the acquisition
Tolman, E. C., Ritchie, B. F., & Kalish, D. (1947). Studies in
of place dispositions. Journal of Comparative and Physiologi-
spatial learning: V. Response versus place learning by the
cal Psychology, 43, 73 – 85.
noncorrection method. Journal of Experimental Psychology,
Salinas, J. A., Packard, M. G., & McGaugh, J. L. (1993). Amygdala
37, 285 – 292.
modulates memory for changes in reward magnitude: Revers-
Viaud, M. D., & White, N. M. (1989). Dissociation of visual and
ible post-training inactivation with lidocaine attenuates the
olfactory conditioning in the neostriatum of rats. Behavioral
response to a reduction in reward. Behavioral Brain Re-Brain Research, 32, 31 – 42. search, 59, 153 – 159.
Weiskrantz, L., and Warrington, E. K. (1979). Conditioning in
Schacter, D. L. (1992). Priming and multiple memory systems:
amnesic patients. Neuropsychologia, 17, 187 – 194.
Perceptual mechanisms of implicit memory. Journal of Cog-
White, N. M. (1989). A functional hypothesis concerning the stria-
nitive Neuroscience, 4, 244 – 256.
tal matrix and patches: Mediation of S-R memory and re-
Scharlock, D. P. (1955). The role of extramaze cues in place and
ward. Life Sciences, 45, 1943 – 1947.
response learning. Journal of Experimental Psychology, 50,
Zola-Morgan, S., Squire, L., & Mishkin, M. (1982). The neuroanat-
omy of amnesia: Amygdala – hippocampus versus temporal
Sutherland, R. J., & Rudy, J. W. (1989). Configural association
stem. Science, 218, 1337 – 1339.
Campos HS . O médico e o cigarro Opinião O médico e o cigarro RESUMO O tabagismo é um dos principais fatores causadores de doença e morte em todo o mundo. Noventa porcento dos casos de câncer do pulmão e de doença pulmonar obstrutiva crônica (DPOC) são causados pelo fumo, que também está diretamente ligado a graves doenças cardiovasculares, bem como a diversos outros prob
 Dorsolateral caudate nucleus (left) and dorsal hippocampus (right) cannulae placements showing the anterior/posterior extent
of needle tip locations at 0.5-mm sections. Caudate nucleus placements ranged from 0.20 to 0.30 mm from bregma. Hippocampalplacements ranged from 02.8 to 03.0 mm from bregma. (Plates adapted from atlas of Paxionos and Watson, 1986).
Dorsolateral caudate nucleus (left) and dorsal hippocampus (right) cannulae placements showing the anterior/posterior extent
of needle tip locations at 0.5-mm sections. Caudate nucleus placements ranged from 0.20 to 0.30 mm from bregma. Hippocampalplacements ranged from 02.8 to 03.0 mm from bregma. (Plates adapted from atlas of Paxionos and Watson, 1986).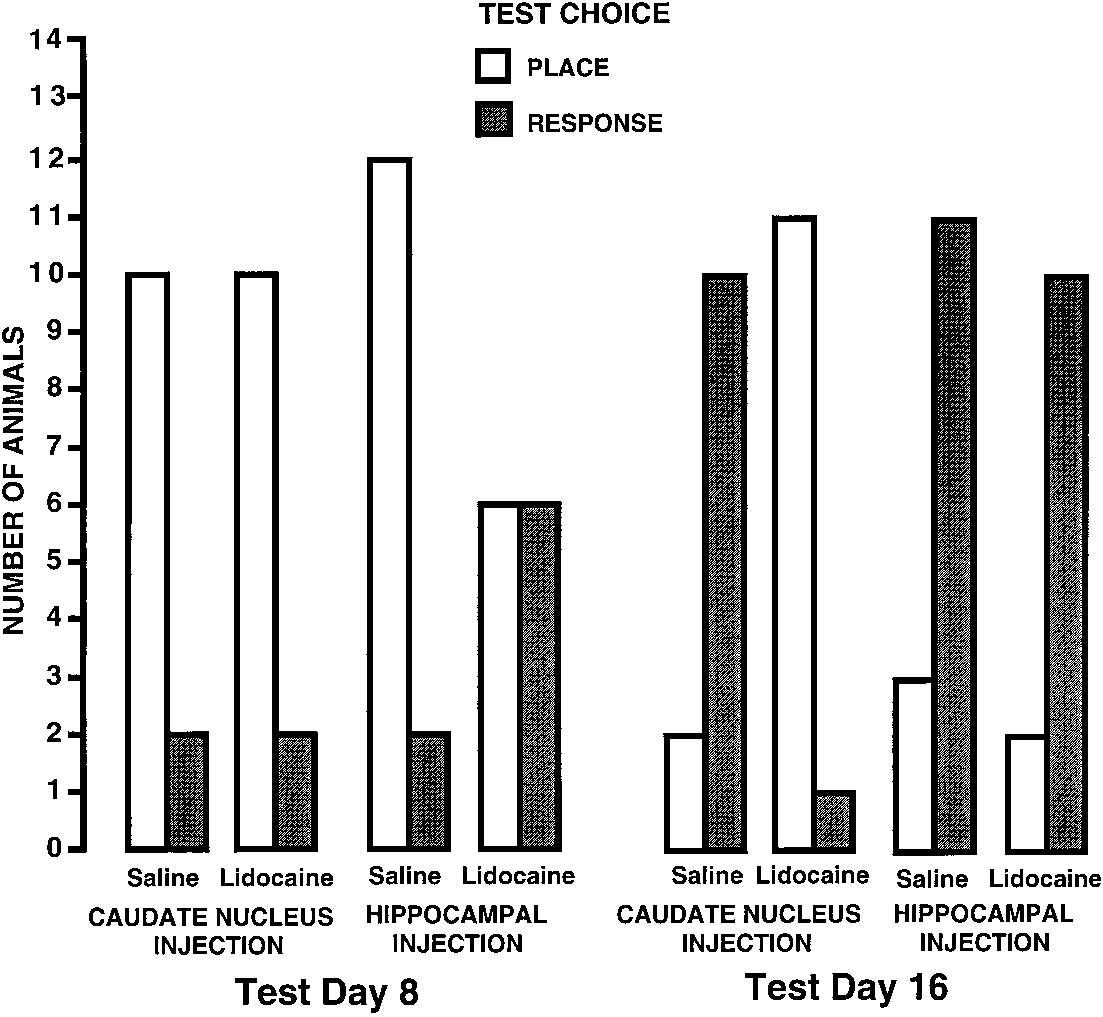 INACTIVATION OF HIPPOCAMPUS OR CAUDATE NUCLEUS
the Day 8 probe trial, animals given saline injectionsinto either the hippocampus or caudate nucleus werepredominantly place learners (hippocampus – salinex2 Å 7.14, p õ .05; caudate nucleus – saline x2 Å5.34, p õ .05). Rats given intracaudate injections oflidocaine were also predominantly place learners (x2Å 5.34, p õ .05), while those receiving intrahippo-campal injections of lidocaine did not show a signifi-cant trend toward either place or response learning(x2 Å 0, n.s.). These findings indicate that functionalinactivation of the hippocampus, but not caudate nu-cleus, prevented the expression of place learning.
INACTIVATION OF HIPPOCAMPUS OR CAUDATE NUCLEUS
the Day 8 probe trial, animals given saline injectionsinto either the hippocampus or caudate nucleus werepredominantly place learners (hippocampus – salinex2 Å 7.14, p õ .05; caudate nucleus – saline x2 Å5.34, p õ .05). Rats given intracaudate injections oflidocaine were also predominantly place learners (x2Å 5.34, p õ .05), while those receiving intrahippo-campal injections of lidocaine did not show a signifi-cant trend toward either place or response learning(x2 Å 0, n.s.). These findings indicate that functionalinactivation of the hippocampus, but not caudate nu-cleus, prevented the expression of place learning.