Kamagra enthält Sildenafilcitrat als pharmakologisch aktiven Bestandteil. Dieser hemmt selektiv die Phosphodiesterase-5 und erhöht dadurch die Konzentration von cGMP im Corpus cavernosum. Der Effekt ist zeitlich begrenzt, da die Halbwertszeit von Sildenafil etwa vier Stunden beträgt. In der galenischen Form als Mundgel erfolgt die Resorption besonders rasch, was zu einem schnelleren Wirkeintritt führt. Der Abbau erfolgt überwiegend hepatisch über CYP3A4, wobei ein aktiver Metabolit entsteht, der zur Gesamtwirkung beiträgt. Typische Nebenwirkungen ergeben sich aus der Vasodilatation, darunter leichte Kopfschmerzen und nasale Kongestion. In klinischen Beschreibungen wird kamagra oral jelly im Zusammenhang mit der schnelleren Absorption erwähnt.
0554.p65
M.R. HEUPEL ET AL. JOURNAL OF EXPERIMENTAL ZOOLOGY 284:586–594 (1999) Plasma Steroid Hormone Profiles and Reproductive Biology of the Epaulette Shark, Hemiscyllium ocellatum
M.R. HEUPEL,* J.M. WHITTIER, AND M.B. BENNETTDepartment of Anatomical Sciences, University of Queensland, St. Lucia,Queensland, Australia 4072
Examination of the reproductive biology of the oviparous epaulette shark,
Hemiscylliumocellatum, was conducted on a wild population. Male sharks were found to reachmaturity at between 55–60 cm total length (TL) and female sharks mature around 55 cm TL. Blood samples collected from mature male and female sharks were analyzed for sex steroid hor-mones to examine seasonal hormone patterns. Plasma samples were analyzed via radioimmu-noassay techniques with female samples measured for estradiol, progesterone, and androgenconcentrations, and male samples measured for androgen concentrations. Male androgen concen-trations showed a single broad peak from July to October with maximum hormone concentrations(60 ng/ml) occurring in August. Male androgen concentrations were lowest in December–February(<20 ng/ml), and appeared to correlate with reproductive activity and water temperature. Femaleandrogen concentrations were an order of magnitude lower than those for males and showed peaksin June (6 ng/ml) and December (8 ng/ml). Estradiol concentrations in females peaked during themonths of September–November (0.5 ng/ml) coinciding with the egg laying period. Progesteroneconcentrations ranged up to 0.5 ng/ml prior to the mating season. Observations of ova size and eggproduction showed eggs develop in pairs and ova are ovulated at a size of 25–27 mm. Females layeggs from August to January. Males were observed with swollen claspers from July through De-cember, with the highest amount of sperm storage in the epididymis occurring between Augustthrough November. Our observations indicate that epaulette sharks in the waters near HeronIsland mate from July through December. J. Exp. Zool. 284:586–594, 1999.
The epaulette shark, Hemiscylliumocellatum,
studied in a range of sharks and rays in order to
is a small benthic shark commonly found in shal-
characterize the reproductive patterns of these
low water on coral reefs in northern Australia and
fishes (e.g., Sumpter and Dodd, ’79; Koob et al.,
New Guinea (Last and Stevens, ’94). Previous re-
’86; Rasmussen and Gruber, ’90, ’93; Callard et
search on aquarium-held individuals showed that
al., ’91, ’93, ’95; Rasmussen and Murru, ’92;
this species bred year-round in a captive environ-
Manire et al., ’95; Manire and Rasmussen, ’97).
ment, producing up to 50 eggs per year (West and
Callard et al. (’91) described the hormone cycles
Carter, ’90), but nothing is known of the repro-
for oviparous and viviparous reproductive strate-
ductive biology of this species in the wild.
gies as either synchronous or asynchronous. The
Defining the breeding cycle of a wild population
oviparous strategy was defined as a synchronous
is an integral component to understanding the bi-
cycle in which estradiol and progesterone concen-
ology of a species. Some elasmobranch species,
trations peak at the same time. Oviparous spe-
such as the lesser spotted dogfish, Scyliorhinus
cies were assumed to have a short cycle in which
canicula, and the black dogfish, Centroscyllium
both hormones increase during the follicular
fabricii, appear to have a continuous breeding sea-
growth phase and decline during the luteal phase.
son (Sumpter and Dodd, ’79; Yano, ’95), whereas
An asynchronous pattern was used to define the
many other species including the bonnetheadshark, Sphyrnatiburo, (Manire et al., ’95), the Aus-tralian sharpnose shark, Rhizoprionodontaylori,
Grant sponsors: The Australian Coral Reef Society; Great Barrier
(Simpfendorfer, ’92) and the blacktip shark, Car-
Reef Marine Parks Authority; University of Queensland, AustraliaPostgraduate Research Scholarship. charhinuslimbatus, (Castro, ’96) have distinct sea-
Work was conducted under QFMA permit numbers 6435 and
PRM000801 and GBRMPA permit numbers G95/454 and G95/595.
*Correspondence to: M.R. Heupel, Mote Marine Lab, 1600 Ken
Thompson Parkway, Sarasota, FL 34236-1096. 1999 WILEY-LISS, INC. REPRODUCTION IN THE EPAULETTE SHARK
viviparous reproductive strategy with estradiol be-
by Maruska et al. (’96): stage 1, primary or germi-
ing dominant and peaking in the follicular growth
nal zone; stage 2, early spermatocysts; stage 3,
phase and progesterone dominant and peaking in
spermatocytes; stage 4, spermatids; stage 5, im-
the luteal phase which occurs later in the cycle.
mature sperm; stage 6, mature spermatocysts; and
Our research into the reproductive biology of
the epaulette shark was designed to examine the
Blood samples for plasma hormone analysis were
reproductive biology of this species and the sea-
taken from five mature male and five mature fe-
sonality of reproductive activity in the wild on a
male sharks for each calendar month. A heparin-
tropical reef off Heron Island, Queensland, Aus-
ized syringe was used to take a 1 ml blood sample
tralia. Components of the study involved anatomi-
from the caudal vessel of sharks. Blood was trans-
cal observation, histological analysis and sex
ferred to an eppendorf tube, stored on ice for up to
steroid hormone analysis to determine if there was
2 hr after which samples were centrifuged, the
a defined breeding season in this species.
plasma pipetted off into clean eppendorf tubes, andstored frozen at –70°C. Plasma samples from male
MATERIALS AND METHODS
sharks were assayed for androgens and samples
Hemiscylliumocellatum were captured during
from females were analyzed for estradiol, progest-
low tides by hand netting over the reef flat area
erone and androgens. Samples were analyzed
on Heron Island Reef. This large platform reef sur-
using coated-tube radioimmunoassay kits for es-
rounds Heron Island, a coral cay situated at
tradiol, progesterone, and androgens (ICN Diag-
23°27'S and 151°55'E. Approximately 500 sharks
nostics, Costa Mesa, CA) and counted using a
were examined during tagging for a mark recap-
gamma counter (Beckman, Fullerton, CA). Coated
ture study. The reproductive condition (i.e., gravid,
tube assay kits were validated by comparing re-
carrying egg purses) and any obvious evidence of
sults of a pooled sample including five female
mating activities of mature females were noted.
samples with results for the same pooled sample
The size (inner clasper length) and condition of
analyzed by traditional radioimmunoassay meth-
claspers of mature males were recorded and cal-
ods. Due to practical considerations the assays
cification of claspers was examined to estimate
were run in two separate batches. Samples from
January, May, and August formed the second
Mature females from all months except May,
batch along with additional samples from other
June, July, and December and mature males from
months. Due to a change in the assay (by the
all months except February, May, June, and July
manufacturer) the values for progesterone and es-
were collected for reproductive organ examination.
tradiol in the second assay were significantly lower
These specimens (32 female and 12 male) were
than in the original assay and therefore these data
used in morphometric as well as histological ex-
amination. Measurements of oviducal glands
Hormone concentrations were analyzed statisti-
(length, width, thickness) and ova diameter were
cally using Sigmastat (Jandel Scientific, San
taken and oviducal gland measurements were mul-
Raphael, CA). Non-parametric Kruskal-Wallace
tiplied to obtain a volume estimate. Lengths and
one-way ANOVA on ranks followed by an all
widths of testes were measured. These measures
pairwise multiple comparison (Dunn’s method)
were compared to the size of the animal, time of
were conducted using a critical probability value
year, and condition of organs (e.g., active, inactive,
of 0.05. Mean hormone concentrations were corre-
regressing). Samples of testes were immersion fixed
lated with mean monthly maximum water tem-
in 4% formaldehyde for subsequent examination.
peratures obtained from records held at Heron
These samples were processed for histology (Sh-
annon Citadel 2000) and embedded in paraffin wax. Sections (7 µm) were cut and mounted on glass
slides before staining with Masson’s trichrome. Males
Stained sections were examined and photographed
Clasper elongation and calcification in male
under light microscopy (Zeiss Axiophot, Germany).
sharks generally occurred when sharks were be-
Testes were cut in cross section and the number of
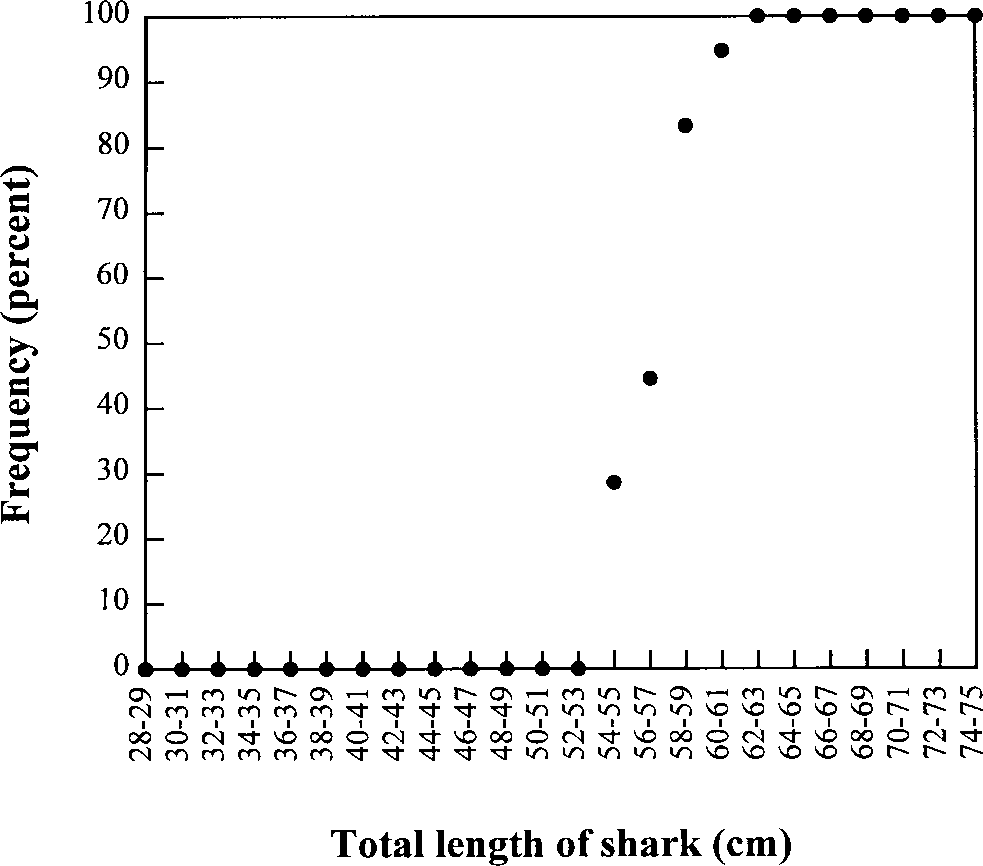
tween 55–60 cm total length (TL) (Fig. 1) with the
spermatocysts in each stage were counted, mea-
smallest mature male 54 cm TL and the largest
sured, and expressed as a percentage of total
immature male 61 cm TL. Inner length measure-
spermatocysts. Individual stages of spermatogen-
ments of fully calcified claspers were consistently
esis were categorized into seven stages as described
about 7% of the total body length of the shark. M.R. HEUPEL ET AL. TABLE 1. Average monthly water temperatures from Heron
stages of spermatogenesis showed the expansion
Island Reef recorded by research station staff1
of spermatocysts from stages 1–5 (Fig. 2). Stage
1 cells were generally about 1.0 µm in diameter. Stage 7 spermatocysts were not measured due to
their degenerative state. Sertoli cell size was con-
sistent throughout the year and ranged from 0.7–
1.0 µm in diameter. When comparing the size of
spermatogenic stages by month it was noted that
on average November samples had the largest
spermatocysts (i.e., stage 5: 34 mm). Samples from
September and December were of similar sizes
(stage 5: 30 mm in both), but specimens from Feb-
ruary were considerably smaller (stage 5: 16 mm).
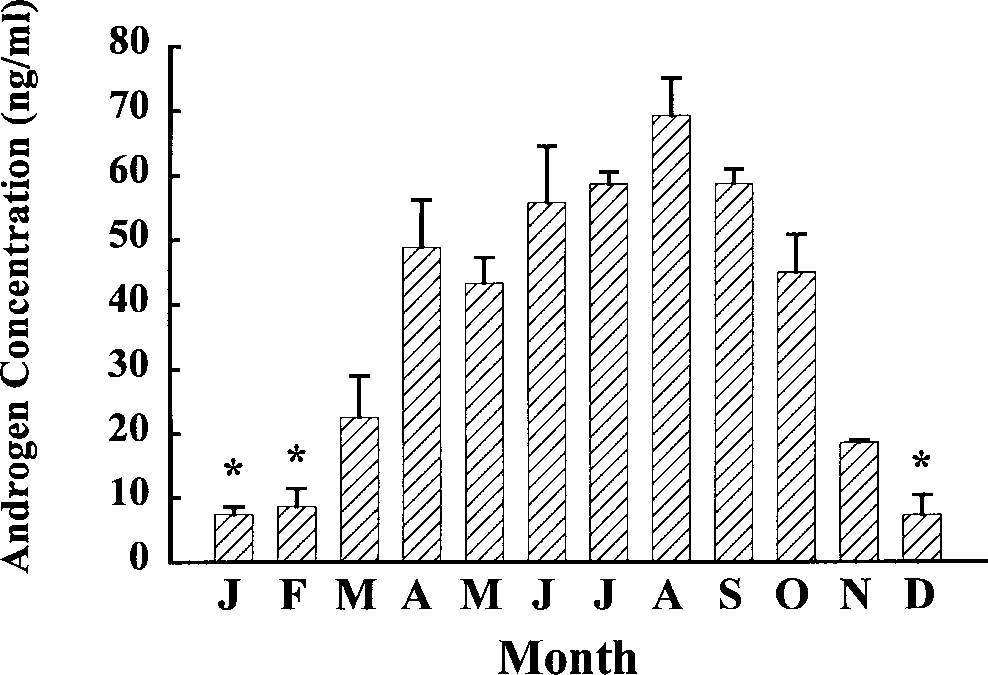
A distinct annual cycle in androgen concentra-
1Water temperature was measured daily from the jetty adjacent to
tions was observed in male sharks (Fig. 3). Dif-
the study site at a depth of 1 m at 8:30 am. Note: the thermometer is
ferences in androgen concentrations between
attached to a float to maintain a depth of 1 m at all times.
months were statistically significant (Kruskal-Wallace = 48.5, P < 0.01). Hormone concentra-
Histological examination of sections of testes
tions were significantly lower (P < 0.05) from
showed the various stages of sperm production
December through February (southern hemi-
throughout the year (Table 2). In April sharks had
sphere summer) with concentrations of <20 ng/
begun sperm production for the mating season.
ml observed. Concentrations rose gradually and
Stages 1–6 were present and 50–75% of sperma-
peaked in July–October at about 60 ng/ml before
tocytes of individual testes were in stages 3 or 4.
starting to decline in November. There was an
In August all stages of sperm production were
inverse correlation between androgen concentra-
present, with a limited portion of the testis de-
tions and water temperature (r2 = 0.93). The high-
voted to stages 1 and 7. The epididymis contained
est concentrations of androgen coincided with
sperm during August–November, with fullness
observations of males with red and swollen clasp-
appearing to be greatest in November. Testes in
ers. Males in this condition were frequently found
this condition were observed to be enlarged com-
between July to December and were assumed to
pared to previous months, and contained about
50% of spermatocysts in stages 5 or 6. Females
Females were found to mature at approximately
55 cm TL. Females less than 55 cm had thin strap-like ovaries and only small non-yolky ova present. Females above this size had well developed ova-ries with yolky ova present.
Measurements of 32 oviducal glands from ma-
ture females showed a change in size through theyear (Fig. 4). Oviducal glands were smallest inJanuary–April. Subsequently, glands showed anincrease in width during August–November whensharks were reproductively active.
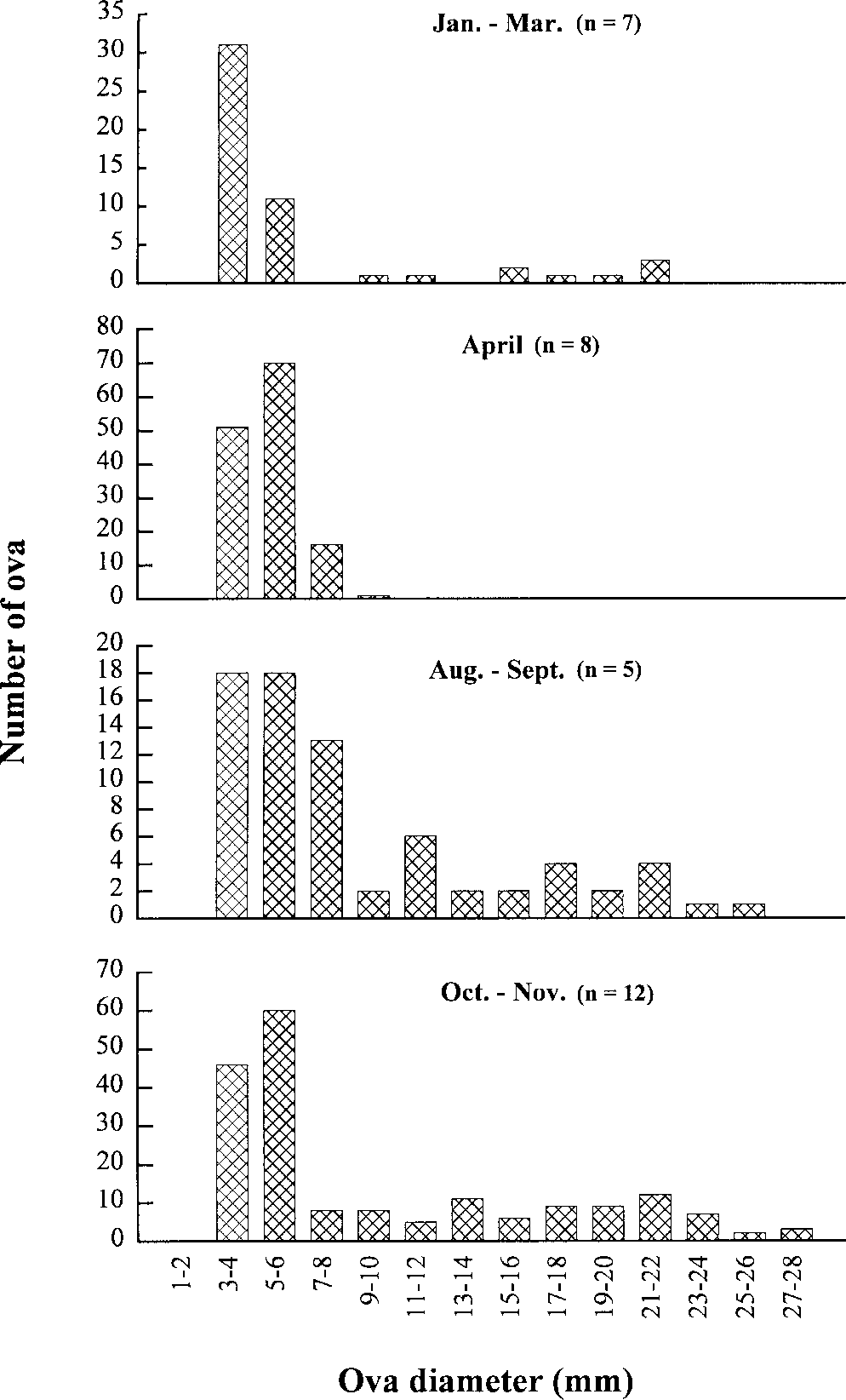
Sizes of vitellogenic ova were variable through-
out the year with a range of 3–27 mm (Fig. 5). Small ova (3–6 mm) were present in all femalesexamined. Females sampled in January had fewlarge ova present, and in February–March femaleshad few ova that appeared to be undergoing re-sorption. By April there were small numbers (10–15 per individual) of yolky ova that were 3–5 mm
Percentage of 249 male Hemiscylliumocellatum
in diameter. In August females had at least five
with fully calcified claspers as a function of total length.
pairs of large, yolky ova of varying sizes. The larg-
REPRODUCTION IN THE EPAULETTE SHARK TABLE 2. Testicular activity of mature male Hemiscyllium ocellatum throughout the year indicating presence of spermand stages of spermatogenesis present
est observed ova were about 25–27 mm with all
35 mm, green-brown in color with fine hair-like
subsequent pairs smaller. The presence of egg pairs
clumps of tendrils that covered the entire surface.
at this stage was observed throughout the remain-
Androgen concentrations measured in females
der of the breeding season (September–November).
were about an order of magnitude lower than
No samples from ovaries were obtained in Decem-
those in males (2–8 ng/ml) (Fig. 6a). Androgen con-
ber, but pairs of egg capsules were collected from
centrations were not significantly different be-
females during August, October, November, Decem-
tween months (Kruskal-Wallace = 11.8, P = 0.38)
ber, and January. Examination of females during
and there was no correlation between water tem-
tagging excursions revealed gravid or pregnant fe-
perature and hormone concentrations (r2 < 0.01).
males from August through early January. Females
Estradiol and progesterone appeared to have sea-
were also noted to have red, irritated tissue around
sonal patterns (Fig. 6b, c). Estradiol concentrations
the cloaca during the months of July and August.
were low during the southern autumn and winter
This was probably a result of mating activities. It
(March–Aug.) with concentrations of 0.05–0.2 ng/
was presumed that ova were ovulated at a size of
ml. Concentrations rose to a peak (0.5 ng/ml) in
25–27 mm since this was the largest size of ova
spring and early summer (September–November)
before declining again in December–February. Es-
Egg capsules were produced in pairs with at
tradiol concentrations were significantly different
least half of the egg capsule formed before ovula-
between months (Kruskal-Wallace = 33.1, P < 0.01)
tion. One female collected for dissection in August
but sample size was not large enough to distin-
had partially developed egg capsules within her
guish where differences occurred. There was a
uterus. The egg capsules were half formed, but
weak inverse correlation between water tempera-
no ovum had been ovulated. There were several
ture and estradiol concentration (r2 = 0.19). Proges-
pairs of large yolky ova present in the ovary sug-
terone concentrations showed a different cycle by
gesting the female was capable of ovulation. Egg
peaking in autumn and winter months (June–July)
capsules at deposition were approximately 90 ×
at concentrations up to 0.5 ng/ml. Concentrationsdecreased slightly in September–October and con-
Distribution of average monthly androgen concen-
Average diameter of spermatocysts in stages 1–6
trations (ng/ml) with standard errors for male H.ocellatum
measured from male H.ocellatum in the month of Novem-
sampled on Heron Island Reef. Asterisks indicate months with
significantly lower (P < 0.05) androgen values. M.R. HEUPEL ET AL.
Yearly volume distribution of oviducal glands from
32 mature female H.ocellatum excluding the months of May,June, July, and December.
tinued this trend through the remainder of theyear. Progesterone concentrations were also sig-nificantly different between months (Kruskal-Wallace = 17.6, P = 0.03) but differences could notbe statistically determined due to restricted samplesizes. There was a slightly stronger inverse corre-lation (r2 = 0.37) to water temperature than thatfor estradiol. DISCUSSION
Information pertaining to the life history of
hemiscyllid sharks in Australian waters is lim-ited. Although these species are commonly ob-served and are generally known to be oviparous(Compagno, ’84; Last and Stevens, ’94), their re-
Measurements of ova diameter from 32 mature
productive timing and periodicity are unknown.
female H.ocellatum throughout the year. Graphs depict the
The limited research available on H.ocellatum in-
progression of ovum size, number, and development throughthe year.
cludes one study on sharks maintained in a con-trolled aquarium environment (West and Carter,’90). There are no data available concerning the
bonnethead shark, S.tiburo (Manire and Ras-
reproductive activities of H.ocellatum in its natu-
mussen, ’97). The claspers of mature R.terrae-novae were found to be about 7–8% of total body
Male and female H.ocellatum were determined
length (Parsons, ’83), similar to the measures
to be reproductively mature at similar sizes. This
found for H.ocellatum in this study.
was based on male clasper calcification and ex-
Histological examination of testes showed that
amination of female reproductive tracts. Using cal-
sperm production had a seasonal cycle. Testes
cification of claspers to determine sexual maturity
were found to be inactive during the months of
in male sharks has been used in many studies on
January–March, a period when androgen concen-
elasmobranch species, including the Atlantic
trations were at their lowest. Sperm production
sharpnose shark, Rhizoprionodonterraenovae
began in April and continued to increase through
(Parsons, ’83), the blue shark, Prionaceglauca
the months of August–November with all stages
(Pratt, ’79), the chain dogfish, Scyliorhinusretifer
of sperm production present. Sperm production
(Castro et al., ’88) the sandbar shark, Carchar-
increased as androgen concentrations also began
hinusplumbeus (Joung and Chen, ’95) and the
to increase. The epididymis contained the larg-
REPRODUCTION IN THE EPAULETTE SHARK
rose prior to the breeding season and remainedhigh throughout the remainder of the egg layingseason. Testosterone concentrations in the lemonshark, Negaprionbrevirostris, and other carchar-hinids were high during the breeding season, butconcentrations vary widely among species. Ne-gaprionbrevirostris had a range of 75–110 ng/mltestosterone, while a study of several species ofcarcharhinid sharks reported a range of 0.85–358ng/ml. Our results from male H.ocellatum fallslightly below concentrations for N.brevirostris,but were within the range of those found for othercarcharhinid species. Based on these results, wepropose that testosterone may be important insexual behaviors, reproductive functions, or mayserve as a precursor for other unidentified steroids(Rasmussen and Gruber, ’90, ’93). Reproductive ac-tivity in male sharks may also be related to watertemperature based on the inverse correlation be-tween water temperature and androgen concentra-tions. The end of the mating season and decreasein androgens coincide with water temperature in-creases during summer months. Whether watertemperature plays any role as a reproductive cuefor male H.ocellatum is unknown, but should beinvestigated further.
Estradiol concentrations vary among species as
well as throughout the reproductive cycle. Stud-ies on various carcharhinid sharks report estra-
Distribution of average monthly (a) androgen, (b)
diol concentrations ranging from 0.4–4.5 ng/ml
estradiol, and (c) progesterone concentrations (ng/ml) with
and 0.6–2.0 ng/ml (Rasmussen and Gruber, ’90;
standard errors for mature female H.ocellatum sampled on
Rasmussen and Murru, ’92). Rajaerinacea had
estradiol concentrations between 0.2–2.0 ng/mldepending on the reproductive status of the fe-
est amount of sperm during November and in De-
male (Koob et al., ’86). Estradiol concentrations
cember sperm production dropped off as the
of the bonnethead shark, S.tiburo, were analyzed
mating season ended. This suggests that male ep-
throughout the reproductive cycle. Concentrations
aulette sharks are producing sperm for mating
were lowest during early pregnancy (mean = 0.20
during the second half of the year. Males were
ng/ml) but increased at mating (mean = 8.98 ng/
observed to have red, swollen claspers from July–
ml) and peaked prior to ovulation (mean = 25.03
November. These observations support the hor-
ng/ml) (Manire et al., ’95). Although estradiol con-
monal data where androgen concentrations were
centrations measured in H.ocellatum appeared
highest from June to October and suggest that
low with a peak of 0.5 ng/ml, these results are
males generally mate between July and Novem-
similar to other oviparous species (e.g., Koob et
ber. Parsons and Grier (’92) defined a seven-stage
al., ’86; Callard et al., ’91). Estradiol concentra-
process of spermatogenesis for S.tiburo and
tions in H.ocellatum increased to their peak dur-
stated that not all spermatogenic stages were
ing the period of egg laying while decreasing and
present throughout the year. Based on their study
remaining low during the period of regression and
Parsons and Grier (’92) concluded that many
shark species may undergo an annual testicular
Increases in estradiol concentrations during the
cycle of regression and recrudescence, while fewer
follicular growth phase are common and have been
species may have spermatogenic stages present
linked to follicle size in the skate, R.erinacea, with
estradiol concentrations increasing in parallel
In H.ocellatum, male androgen concentrations
with follicle size (Koob et al., ’86). Further re-
M.R. HEUPEL ET AL.
search on R.erinacea and Squalusacanthias sup-
tion season. Manire et al. (’95) reported progest-
ported these data and showed that increases in
erone increased during preovulation (mean = 8.9
both estradiol and testosterone characterized the
ng/ml) and ovulation (mean = 16.6 ng/ml) prior to
follicular phase (Callard et al., ’93). Research on
a peak after ovulation (mean = 26.6 ng/ml) in S.
carcharhinid sharks also showed an increase in
tiburo. This result is similar to that described for
estradiol just prior to mating as oocytes were ma-
the dogfish S.acanthias (Callard et al., ’93). How-
turing (Rasmussen and Gruber, ’90, ’93; Ras-
ever, both S.tiburo and S.acanthias are vivipa-
mussen and Murru, ’92). This increase is thought
rous species and show a different pattern from
to set ovulatory events in motion or may regulate
the one described for the oviparous skate R.
the reproductive cycle (Rasmussen and Murru, ’92;
erinacea. Koob et al. (’86) reported an elevation
Rasmussen and Gruber, ’93). Estradiol concentra-
in progesterone for a restricted two day period be-
tions in female H.ocellatum were highest in the
fore encapsulation with a sharp drop on the day
second half of the year and would coincide with
of encapsulation and low concentrations through-
maximum ova sizes, ovulation and egg laying.
out the rest of the year. Callard et al. (’93) re-
Fluctuations in estradiol concentrations were cor-
ported elevated concentrations of progesterone
related with changes in water temperature, but
pre- and peri-ovulation in R.erinacea. Serial
because the relationship was weak it is unlikely
samples examined from one captive female H.
that water temperature plays a role in the timing
ocellatum showed a peak in progesterone the
morning the eggs had been laid (Heupel, unpub-
Androgen concentrations in females of other spe-
lished data). Concentrations previous to and af-
cies are generally found to parallel estradiol con-
ter this point were essentially undetectable,
centrations. Several species have been analyzed
suggesting that progesterone is most active at ovi-
and showed an increase in testosterone concen-
position in H.ocellatum. Progesterone is thought
trations (along with estradiol) prior to and dur-
to regulate events associated with ovulation, en-
ing mating (Callard et al., ’91; Rasmussen and
capsulation, and egg retention in oviparous spe-
Murru, ’92; Rasmussen and Gruber, ’93; Manire
cies and may have specific triggering roles in
et al., ’95). Koob et al. (’86) found testosterone fluc-
viviparous species. Progesterone may also inhibit
tuated with estradiol, but was present in higher
activities such as vitellogenesis (Koob et al., ’86;
concentrations. Testosterone concentrations de-
Rasmussen and Murru, ’92; Callard et al., ’93;
crease after mating and remain low throughout
the rest of the reproductive cycle of many elas-
It has been suggested that hormone concentra-
mobranch species (Rasmussen and Murru, ’92;
tions in females of oviparous species peak more
Rasmussen and Gruber, ’93). Testosterone may be
than once during a season (Koob et al., ’86; Callard
important in initiating some sequential ovulatory
et al., ’91, ’93, ’95). However, this was not the case
events and may have a role in courtship. Due to
in our studies of H.ocellatum, and was not seen
its lower concentrations throughout the rest of the
in several other studies on oviparous species. Re-
cycle, it does not appear to be important during
search by Sumpter and Dodd (’79) examined the
pregnancy in viviparous species (Rasmussen and
hormone cycles of the lesser spotted dogfish, S.
Murru, ’92; Rasmussen and Gruber, ’93). Andro-
canicula. This species is oviparous and has an ex-
gen concentrations in female H.ocellatum did not
tended, if not continuous, breeding season. Despite
vary significantly throughout the reproductive sea-
the extended reproductive cycle of this species, es-
son. Although it is possible that androgens are
tradiol and testosterone concentrations displayed
important in initiating changes in the reproduc-
a distinct annual cycle. Both hormones fluctuated
tive tract, no supporting evidence was found from
together, rising as the ovary recrudesced and fall-
androgen levels in H.ocellatum.
ing as the rate of egg laying decreased. Although
Progesterone concentrations in female H.ocell-
this study did not include progesterone analysis
atum were usually low except for a peak from
it clearly defined one estradiol peak rather than
April–July prior to the egg laying period. As with
several throughout the season. This pattern of one
estradiol, there was a weak correlation between
single peak in hormone concentrations per year
water temperature and hormone concentrations,
is similar to that observed for H.ocellatum
but water temperature probably does not play a
major role in progesterone activity. Peaks in
Most oviparous elasmobranchs produce eggs in
progesterone concentrations are thought to help
pairs (Luer and Gilbert, ’85; Castro et al., ’88; Ellis
prepare the reproductive tract for the egg produc-
and Shackley, ’95; Yano, ’95). Epaulette sharks also
REPRODUCTION IN THE EPAULETTE SHARK
produce eggs in pairs and appear to ovulate ova
egg pairs is variable among oviparous elasmo-
into egg capsules after they are at least half
branch species. The thornback ray, Rajaclavata,
formed. Studies of at least two other oviparous
can produce a pair of eggs from 0–2 days after
elasmobranchs, the dogfish S.canicula, and the
the previous pair (Ellis and Shackley, ’95). The
clearnose skate, R.eglanteria, have shown simi-
clearnose skate, R.eglanteria, takes slightly
lar patterns of ovulation. Ova were not present
longer with 4.5 ± 2.2 days between egg pairs (Luer
in egg capsules less than three-fourths formed in
and Gilbert, ’85) and the chain dogfish, S.retifer,S.canicula (Metten, ’39) and R.eglanteria formed
requires 14–16 days between laying egg pairs
two-thirds of the egg capsule prior to ovulation
(Castro et al., ’88). The period between egg pair
and fertilization (Luer and Gilbert, ’85). Metten
production for H.ocellatum was not determined
(’39) also described one pair of egg capsules that
in the present study. Observation of captive (wild
were fully formed but smaller than normal and
caught) females at Heron Island Reef showed that
without ova. No explanation was given for eggs
none produced more than one pair of eggs (Heupel,
in this condition, and no explanation is obvious
unpublished data). This may have resulted from
for the same condition observed in H.ocellatum
females being kept in isolation when found to be
gravid. One female kept isolated from male sharks
The egg laying behavior of dogfishes has been
produced a pair of empty egg capsules. Whether
well documented with detailed descriptions of at-
this was the result of not having a male present
tachment of the long tendrils of the egg capsule
in the tank, or was due to some other influence,
to a vertical structure and the use of this struc-
is unknown. However, the presence of red, swol-
ture to pull the egg capsule from the oviduct
len claspers and sperm production from July to
(Castro et al., ’88). However, due to the difference
December suggests that males are capable of mat-
in tendrils found on egg capsules of H.ocellatum,
ing throughout the egg laying season.
it is unlikely that they use this type of strategy.
Wourms (’77) defined three types of reproductive
The long hair-like tendrils would appear to be
cycle in elasmobranchs: (1) breeding throughout the
more suited for egg laying similar to that de-
year; (2) partially defined annual cycle with one or
scribed for the clearnose skate, R.eglanteria. The
two peaks during the year; and (3) a well defined
egg laying behavior of R.eglanteria described by
annual or biennial cycle. Although epaulette sharks
Luer and Gilbert (’85) included the female set-
held in a captive aquarium environment fell into
tling quietly on the sediment before contracting
the first category of Wourms’ description (West and
the pelvic fins ventrally, shaking the pelvis from
Carter, ’90), animals sampled in the natural envi-
side to side, and swimming away leaving a single
ronment fell into the last category. The differences
egg capsule on the sediment. This activity was
in results between aquarium-held sharks and wild-
violent enough to leave the egg capsule covered
caught sharks may be due to a lack of seasonal tem-
in sediment from the bottom of the tank. This type
perature variation in the aquarium environment.
of egg laying behavior would appear to be effec-
As shown by correlation of water temperature and
tive for depositing eggs under and among coral,
male testosterone concentrations, seasonal tempera-
and because the egg capsules of H.ocellatum lack
ture changes may be a cue for commencement and
long tendrils, it is likely that this type of method
conclusion of the mating period. Removal of this
would be used to attach egg capsules to coral. One
cue may result in continuous mating activities. Fur-
female H.ocellatum was held in a large aquarium
ther examination of the effects of water tempera-
with a number of different types of shelter and
coral including one small piece of Acropora coral. Although there were several other types of coral
ACKNOWLEDGMENTS
present, the shark placed both egg capsules on
We thank the staff at Heron Island Research
the one piece of Acropora. We were unable to re-
Station for their help throughout this research;
move the egg from the coral by gently pulling the
A. Chan for technical assistance, K. Townsend,
two apart. Although no egg capsules have been
T. Turner, and S. Bennett for field assistance.
discovered on the reef flat at Heron Island Reef,
We also thank Dr. C. Manire and two anonymous
the habits of this species suggest eggs are depos-
reviewers for their advice and comments. This
ited under coral heads. The presence of very small
work was undertaken while the primary author
juvenile sharks in Acropora beds suggest they may
was in receipt of an Overseas Postgraduate Re-
search Scholarship at the University of Queens-
The length of time between laying of successive
M.R. HEUPEL ET AL. LITERATURE CITED
female bonnethead shark, Sphyrnatiburo. Gen CompEndocrinol 97:366–376.
Callard IP, Etheridge K, Giannoukos G, Lamb T, Perez L.
Maruska KP, Cowie EG, Tricas TC. 1996. Periodic gonadal
1991. The role of steroids in reproduction in female elas-
activity and protracted mating in elasmobranch fishes. J
mobranchs and reptiles. Ster Biochem Mol Biol 40:571–575.
Callard IP, Fileti LA, Koob TJ. 1993. Ovarian steroid synthe-
Metten H. 1939. Studies on the reproduction of the dogfish.
sis and the hormonal control of the elasmobranch repro-
Phil Trans Roy Soc Ser B 230:217–241.
ductive tract. Envir Biol Fishes 38:175–185.
Parsons GR. 1983. The reproductive biology of the Atlantic
Callard IP, Putz O, Paolucci M, Koob TJ. 1995. Elasmobranch
sharpnose shark Rhizoprionodonterraenovae (Richardson).
reproductive life histories: endocrine correlates and evolu-
tion. In: Goetz FW, Thomas P, editors. Proceedings of the
Parsons GR, Grier HJ. 1992. Seasonal changes in shark
Fifth International Symposium on the Reproductive Biol-
testicular structure and spermatogenesis. J Exp Zool 261:
ogy of Fish. Austin, TX, 2–8 July. p 204–208.
Castro JI. 1996. Biology of the blacktip shark, Carcharhinus
Pratt HL. 1979. Reproduction in the blue shark, Prionacelimbatus, off the southeastern United States. Bull Mar Sci
glauca. Fishery Bull 77:445–470.
Rasmussen LEL, Gruber SH. 1990. Serum concentrations of
Castro JI, Bubucis PM, Overstrom NA. 1988. The reproduc-
circulating steroid hormones in free-ranging carcharhinoid
tive biology of the chain dogfish Scyliorhinusretifer. Copeia
Rasmussen LEL, Murru FL. 1992. Long-term studies of se-
Compagno LJV. 1984. FAO species catalogue, volume 4. Sharks
rum concentrations of reproductively related steroid hor-
of the world: an annotated and illustrated catalogue of shark
mones in individual captive carcharhinids. Aust J Mar
species known to date. Part 1. Hexanchiformes to Lamni-
formes. FAO Fisheries Synopsis No. 125, Rome. p 188–199.
Rasmussen LEL, Gruber SH. 1993. Serum concentrations of
Ellis JR, Shackley SE. 1995. Observations on egg-laying in
reproductively-related circulating steroid hormones in the
the thornback ray. J Fish Biol 46:903–904.
free-ranging lemon shark, Negaprionbrevirostris. Env Biol
Joung SJ, Chen CT. 1995. Reproduction in the sandbar shark,
Carcharhinusplumbeus, in the waters off northeastern Tai-
Simpfendorfer CA. 1992. Reproductive strategy of the Aus-
tralian sharpnose shark, Rhizoprionodontaylori (Elasmo-
Koob TJ, Tsang P, Callard IP. 1986. Plasma estradiol, test-
branchii: Carcharhinidae), from Cleveland Bay, northern
osterone, and progesterone concentrations during the ovu-
Queensland. Aust J Mar Freshwater Res 43:67–75.
latory cycle of the skate (Rajaerinacea). Biol Reprod
Sumpter JP, Dodd JM. 1979. The annual reproductive cycle
of the female lesser spotted dogfish, Scyliorhinuscanicula
Last PR, Stevens JD. 1994. Sharks and rays of Australia.
L., and its endocrine control. J Fish Biol 15:687–695.
West JG, Carter S. 1990. Observations on the development
Luer CA, Gilbert PW. 1985. Mating behavior, egg deposition,
and growth of the epaulette shark Hemiscylliumocell-
incubation period, and hatching in the clearnose skate, Rajaatum (Bonnaterre) in captivity. J Aquariculture Aquat Sci
eglanteria. Env Biol Fishes 13:161–171.
Manire CA, Rasmussen LEL. 1997. Serum concentrations of
Wourms JP. 1977. Reproduction and development in chon-
steroid hormones in the mature male bonnethead shark,
drichthyan fishes. Am Zool 17:379–410. Sphyrnatiburo. Gen Comp Endocrinol 107:414–420.
Yano K. 1995. Reproductive biology of the black dogfish
Manire CA, Rasmussen LEL, Hess DL, Hueter RE. 1995. Centroscylliumfabricii, collected from waters of western
Serum steroid hormones and the reproductive cycle of the
Greenland. J Mar Biol Assoc UK 75:285–310.
THE ACADEMY OF THE HOLY CROSS INGREDIENT LIST JANUARY 2014 B R O C C O L I C H E D D A R S O U P (G F ) Water, Broccoli, Cheddar Cheese (milk, cheese cultures, salt, enzymes, annatto), Rice Flour, Light Cream, Olive Oil, Butter (cream, salt) Rice Starch, Onions, Sea Salt, Spices C ontains m ilk Bow Tie Pasta, Marinara Sauce (Diced Tomatoes, Tomato Puree, On
Bedienungsanleitung Operating instructions Mode d’emploi Istruzioni per I’uso Instrucciones de manejo Gebruiksaanwijzing Bruksanvisning Was Sie über Ihren Spritzapparat wissen sollten Alle HANSA Spritzapparate sind Präzisions- geräte. Um optimale Spritzergebnisse zu erzielen, ist eine sachgemäße Behandlung erforderlich. Niemals eine Reparatur beginnen, von der Sie
 M.R. HEUPEL ET AL.
M.R. HEUPEL ET AL.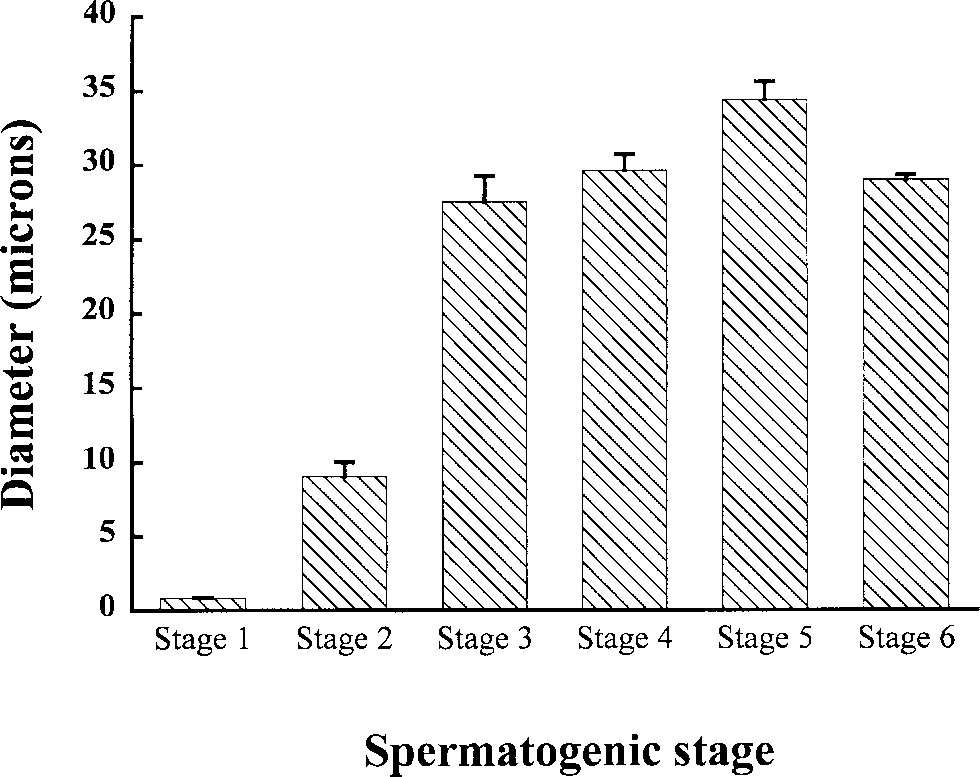
 REPRODUCTION IN THE EPAULETTE SHARK
REPRODUCTION IN THE EPAULETTE SHARK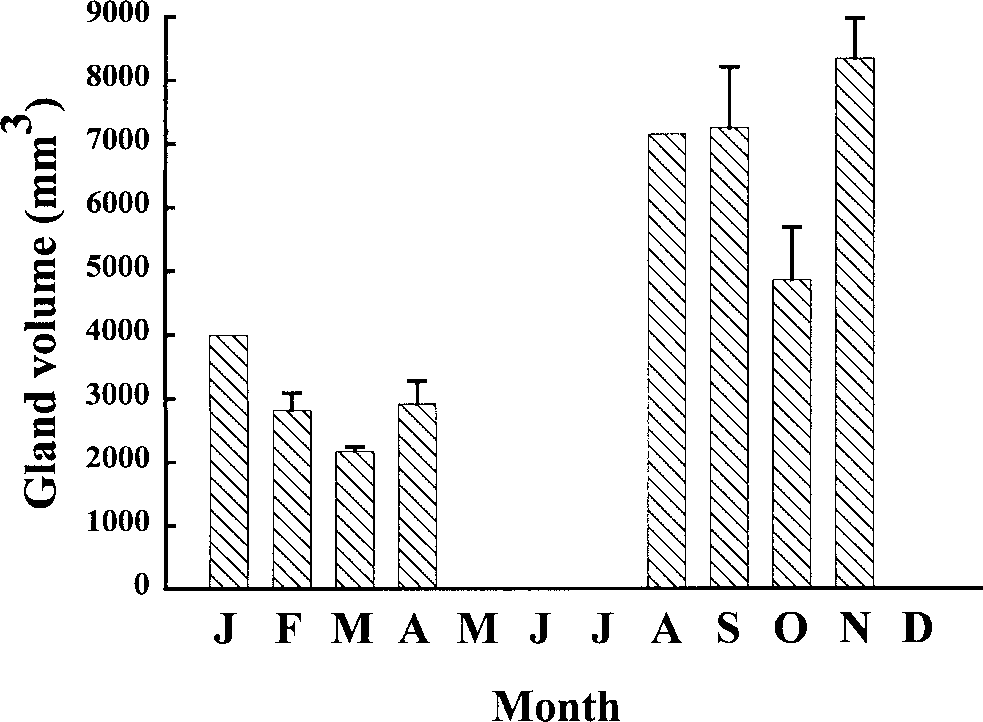
 M.R. HEUPEL ET AL.
M.R. HEUPEL ET AL.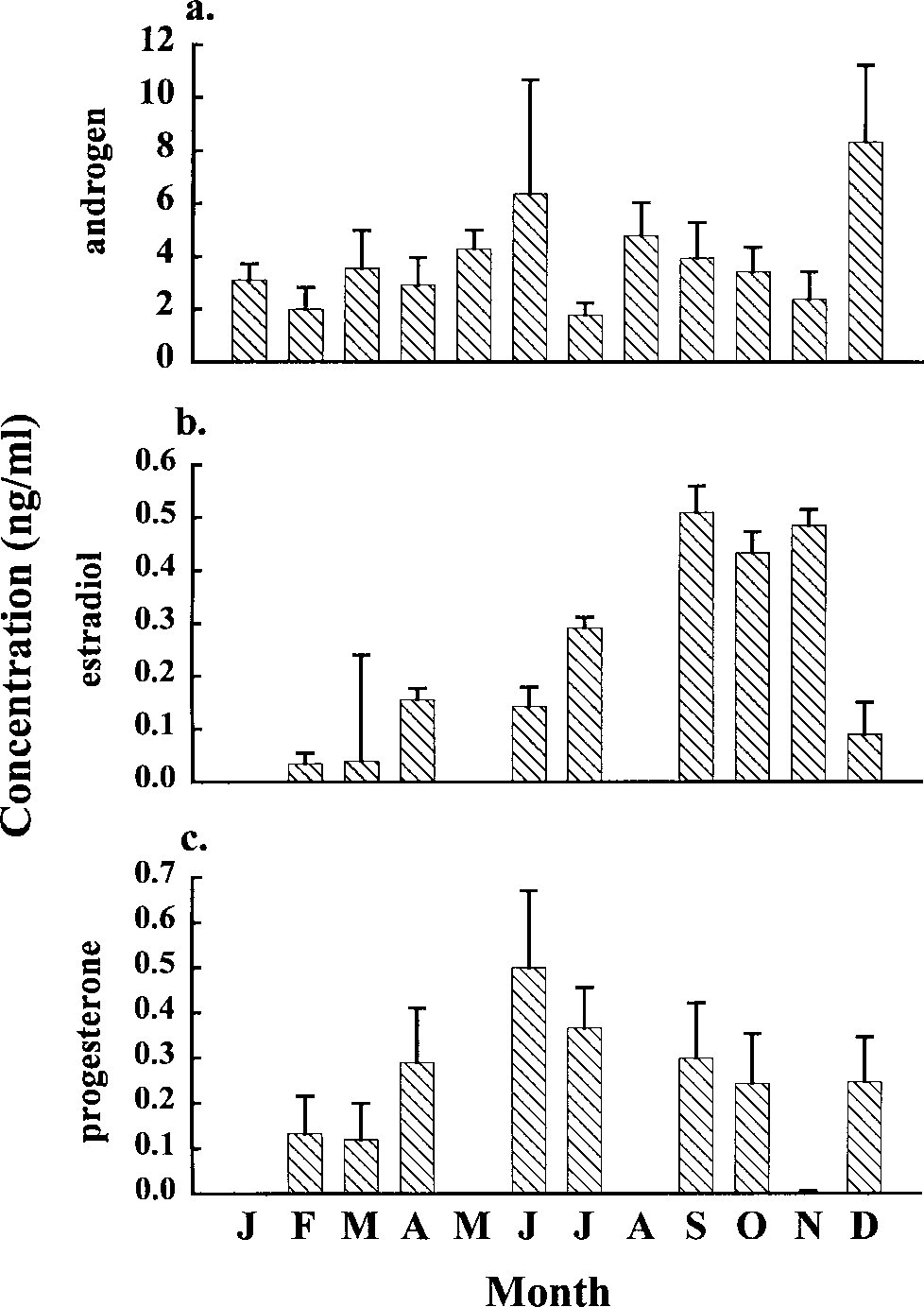 REPRODUCTION IN THE EPAULETTE SHARK
REPRODUCTION IN THE EPAULETTE SHARK